Нестандартная терапия психических и неврологических расстройств ( по материалам статьи Detlev Boison, 2011)

Эта заметка посвящена нуклеозидам: аденозину, гуанозину, цитидину, уридину и инозину, ингибиторам их метаболизма и основанным на нуклеозидах терапевтическим подходам лечения заболеваний головного мозга
Аденозин
В центральной нервной системе деградация АТФ является основным источником синаптического аденозина, тогда как синтез de novo играет лишь вспомогательную роль. В мозге основным путем удаления аденозина является его фосфорилирование до АМФ с помощью аденозинкиназы; альтернативно, аденозин может быть дефосфорилирован аденозин-деаминазой в инозин. Аденозин является повсеместным конечным продуктом реакций трансметилирования. Однако когда уровни аденозина повышаются, равновесие реакции гидролазы s-аденозилгомоцистеина (SAH) смещается в сторону синтеза SAH, что приводит к ингибированию реакций трансметилирования.
Противоположные активности A 1 и A 2A R подразумевают, что аденозин способен реализовать как тормозящие, так и возбуждающие функции в головном мозге. A 1 Rs опосредует ингибирующую нейромодуляцию, связываясь с ингибирующими G i или G o содержащими G белки. В результате этого взаимодействия стимулируется аденилатциклаза, активируются внутренне выпрямляющие K + каналы, а пресинаптические Ca 2+ каналы ингибируются и фосфолипаза С становится активированной. Следовательно, высвобождение преимущественно возбуждающих нейротрансмиттеров, таких как дофамин, серотонин и ацетилхолин, ингибируется, в то время как постсинаптические нейроны остаются гиперполяризованы. Таким образом, терапевтическая активация A 1 Rs приводит к противоэпилептическим, антипсихотическим и нейропротективным эффектам.
Гуанозин
Несмотря на то, что рецепторы и молекулярные пути, на которые воздействует гуанозин, до сих пор не полностью изучены, некоторые исследователи предполагают, что гуанозин является потенциально новой мишенью для нейропротекции и нейромодуляции.
Цитидин и уридин
Цитидин и уридин не оказывают прямого влияния на функцию ЦНС. Через утилизацию пиримидина они трансформируются в цитидин-5′-трифосфат (CTP) и уридин-5′-трифосфат (UTP) и способны вносить свой вклад в синтез фосфатидилхолина и фосфатидилэтаноламина в мозге по пути Кеннеди. На основании этих метаболических связей метаболизм цитидина и уридина может, однако, играть роль в патогенезе нейродегенеративных расстройств.
Инозин
Нуклеозиды в патогенезе нейропсихиатрических расстройств
Эпилепсия
Инсульт
Инсомния
Боль
Про- и антиноцицептивная активность могут быть опосредованы аденозином в зависимости от места применения или вовлеченного рецептора (ов). Патогенез боли с участием A 1 R, который доминирует в опосредовании антиноцицептивных эффектов аденозина, привлекают наибольшее внимание исследователей. Таким образом, агонисты A 1 R, действующие на опаределенные участки позвоночника, являются эффективными антиноцицептивными агентами в условиях невропатической и воспалительной боли, терапевтические эффекты которых, вероятно, опосредованы прединклюзивным ингибированием высвобождения трансмиттера в болевых путях. Аналогичным образом, внутривенное введение аденозина привело к значительному уменьшению нейропатической боли в двойном слепом плацебо-контролируемом перекрестном исследовании I фазы.
Шизофрения
Пуриновые и пиримидиновые основания. нуклеозиды. нуклеотиды

ПУРИНОВЫЕ И ПИРИМИДИНОВЫЕ ОСНОВАНИЯ. НУКЛЕОЗИДЫ. НУКЛЕОТИДЫ
Цель : сформировать знания строения и свойств нуклеотидов, их роли в метаболизме; первичной и вторичной структуры нуклеиновых кислот.
[1] С. 420–444, [2] С. 109–115.
Нуклеиновые кислоты обеспечивают хранение и передачу наследственной информации, непосредственно участвуют в механизмах реализации этой информации путем программированного синтеза всех клеточных белков. Нуклеиновые кислоты представляют собой высокомолекулярные соединения, молекулярная масса которых колеблется в пределах от 25 тыс. до 1 млн дальтон и более. Полимерные цепи нуклеиновых кислот построены из мономерных единиц — нуклеотидов, в связи с чем нуклеиновые кислоты называют полинуклеотидами. Существуют два типа нуклеиновых кислот — ДНК и РНК, различающиеся по молекулярной массе, составу азотистых оснований, сахаров, устойчивости и функциям.
Нуклеиновые кислоты обладают выраженными кислотными свойствами (обусловленными наличием остатков ортофосфорной кислоты в их составе) и при физиологических значениях рН несут отрицательный заряд. Этим объясняется одно из важных свойств нуклеиновых кислот — взаимодействовать с основными белками (гистонами). Нуклеиновые кислоты также образуют ионные связи с катионами металлов, преимущественно с Мg2+.
При полном гидролизе нуклеиновых кислот (нагревании в присутствии хлорной кислоты) в гидролизате обнаруживаются пуриновые и пиримидиновые основания, углеводы (рибоза и дезоксирибоза) и фосфорная кислота:
Дезоксирибоза
Тимин
Рибоза
Урацил
НУКЛЕИНОВЫЕ ОСНОВАНИЯ, ИХ ТАУТОМЕРНЫЕ ФОРМЫ



К пуриновым основаниям относятся аденин и гуанин. Для аденина характерна амино-иминная таутомерия. А для гуанина — лактам-лактимная и амино-иминная.


При физиологических условиях нуклеиновые основания существуют только в лактамной и аминной формах.
Нуклеиновые основания растворимы в воде, их растворы имеют слабощелочную
реакцию среды (рН 8–9).
Цитозин + Рибоза ® Цитидин
Цитозин + Дезоксирибоза ® Дезоксицитидин
Аденин + Рибоза ® Аденозин
Аденин + Дезоксирибоза ® Дезоксиаденозин
Нуклеотиды — это фосфорные эфиры нуклеозидов, являющиеся структурными единицами нуклеиновых кислот. Они образуются в реакции фосфорилирования, протекающей по типу нуклеофильного замещения, приводящей к формированию сложноэфирной связи.
Нуклеотиды способны гидролизоваться в кислой и щелочной среде.

Гидролизу могут подвергаться как N-гликозидная, так и сложноэфирная связи, и в зависимости от рН среды могут образовываться или нуклеозиды или компоненты нуклеотида.
Обнаружить в продуктах гидролиза нуклеотидов пуриновые основания можно при помощи «серебряной пробы».

В ходе реакции образуется светло-коричневый осадок серебряных солей пуриновых оснований.
Пентозы обнаруживают с помощью реакции Биаля.

Фурфурол далее конденсируется с орцином, о чем свидетельствует появление сине-зеленой окраски.
Фосфорную кислоту можно обнаружить при помощи молибденового реактива.
H3PO4 + 12(NH4)2MoO4 + 21HNO3 → (NH4)3PO4 · 12MoO3 + 21NH4 NO3 + 12H2O
Образующийся фосфорномолиденовокислый аммоний — осадок желтого цвета.
Нуклеотиды входят в состав нуклеиновых кислот; но, кроме того, они встречаются
в клетке в свободном состоянии, выполняя энергетическую и регуляторную функции. К наиболее важным из них относятся циклические нуклеотиды, аденозинтрифосфат и гуанозинтрифосфат.


Они содержат две ангидридные связи, называемые макроэргическими. При расщеплении макроэргической связи Р
О (обозначаемой волнистой линией) выделяется
32 кДж/моль. С этим связана важнейшая роль АТФ как «поставщика» энергии во всех живых клетках. Нуклеозидная часть молекулы важна для узнавания и связывания с различными ферментами, использующими АТФ или ГТФ.
При физиологических значениях рН АТФ находится в ионизированном состоянии и в клетке связывается с ионами двухвалентных металлов (Мg2+, Са2+). Такое связывание частично нейтрализует общий отрицательный заряд и облегчает гидролиз АТФ под действием нуклеофилов (ОН-, Н2О). АТФ способен переносить потенциальную энергию на множество важных биологических соединений. Так, с участием АТФ осуществляется активный транспорт ионов через биологические мембраны, активирование аминокислот перед их связыванием с т-РНК, синтез полинуклеотидных цепей, образование пептидных связей в белках и т. д.

Наиболее важными представителями этой группы соединений являются никотинамидадениндинуклеотид (НАД) и его фосфат (НАДФ). Эти соединения выполняют роль коферментов большого числа ферментов дегидрогеназ и, следовательно, являются участниками окислительно-восстановительных реакций. В соответствии с этим они могут существовать как в окисленной (НАД+, НАДФ+), так и восстановленной (НАДН, НАДФН) формах.
Структурным фрагментом НАД+, НАДФ+ является никотинамидный остаток в виде пиридиниевого катиона. В организме человека с участием НАД+ происходит окисление гидроксилсодержащих соединений, например, этанола в ацетальдегид.


Ароматический пиридиниевый цикл в НАД+ в результате окисления гидроксилсодержащего субстрата восстанавливается и переходит в менее стабильное неароматическое
состояние в восстановленной форме НАДН, т. е. энергия НАДН будет выше, чем у НАД+. Повышение энергии в молекуле НАДН происходит за счет части энергии, выделяющейся
в результате окисления молочной кислоты. Таким образом НАДН запасает энергию, которая затем расходуется в других биохимических процессах, требующих энергетических затрат.
ПЕРВИЧНАЯ СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ
Нуклеиновые кислоты представляют собой макромолекулы, построенные из нуклеотидов, соединенных в линейную последовательность фосфодиэфирными связями. Рибонуклеотиды образуют РНК, дезоксирибонуклеотиды — ДНК.
Мононуклеотиды в структуре нуклеиновых кислот связываются друг с другом через остатки фосфорной кислоты, которые образуют две сложноэфирные (фосфодиэфирные)
связи: с С3′ предыдущего нуклеотидного звена и с С5′ последующего нуклеотидного звена. Полимерная цепь нуклеиновых кислот состоит из чередующихся пентозных и фосфатных остатков, а гетероциклические основания являются «боковыми» группами, присоединенными к пентозным остаткам. Концы линейной полинуклеотидной цепи обозначают: 5′-конец (слева) и 3′ — конец (справа). Написание цепи обычно начинают с 5′-конца. В этом случае общее направление образования фосфодиэфирных связей в цепи обозначается 5’→3′. На
5′-конце находится фосфатная группа, и такой конец цепи сокращенно обозначают буквой «Р». На другом конце цепи в пентозном остатке сохраняется свободной гидроксильная группа у С-3′, и поэтому этот конец цепи обозначают как ОН-конец. На рисунке приведено строение участка цепи ДНК, включающего четыре нуклеотида.
Принцип построения цепи РНК такой же, как и у ДНК, с двумя исключениями: пентозным остатком в РНК является D-рибоза, и в наборе гетероциклических оснований используется не тимин, а урацил. Таким образом, первичная структура нуклеиновых кислот — это линейная последовательность нуклеотидных звеньев, связанных ковалентными фосфодиэфирными связями в непрерывную цепь полинуклеотида.

ПОНЯТИЕ О ДВОЙНОЙ СПИРАЛИ ДНК
Вторичная структура ДНК — это пространственная организация полинуклеотидных цепей в ее молекуле. Согласно модели Дж. Уотсона и Ф. Крика, молекула ДНК состоит из двух полинуклеотидных цепей, правозакрученных вокруг общей оси с образованием двойной спирали, имеющей диаметр 1,8–2,0 нм. Эти две полинуклеотидные цепи антипараллельны друг другу, т. е. направления образования фосфодиэфирных связей в них противоположны: в одной цепи 5’–3′, в другой 3’–5′.
Стэкинг-взаимодействия между основаниями стабилизируют спиральную структуру, преодолевая силы электростатического отталкивания между отрицательно заряженными фосфатными группами. Эта энергия стабилизации может быть равна или превышать энергию связывания цепей водородными связями. Построение молекулярных моделей показывает, что именно в правой спирали взаимодействие между нуклеотидами оптимально.
Водородные связи между комплементарными основаниями — это один из видов взаимодействия, стабилизирующих двойную спираль. Данный вид взаимодействия называют «поперечным» в отличие от «вертикального» (стэкинг-взаимодействия). Две цепи ДНК, образующие двойную спираль, не идентичны, но комплементарны друг другу. Это означает, что первичная структура, т. е. нуклеотидная последовательность, одной цепи предопределяет первичную структуру второй цепи.
Пуриновые и пиримидиновые основания нуклеотидных звеньев направлены внутрь двойной спирали. Между пуриновым основанием со стороны одной цепи и пиримидиновым основанием со стороны другой цепи образуются водородные связи. Эти основания, связанные водородными связями составляют комплементарные пары.

 Комплементарность цепей составляет химическую основу важнейших функций ДНК — хранения и передачи наследственных признаков. При делении клеток двойная спираль ДНК раскручивается и разделяется на две части. На каждой отдельной цепи, как на матрице, происходит биосинтез новой цепи ДНК с учетом принципа комплементарности. Вновь образовавшаяся цепь не идентична, но комплементарна исходной матрице. В результате воссоздаются две новые двойные спирали ДНК. Такой процесс называется репликацией, и он лежит в основе обеспечения дочерних клеток молекулами ДНК, идентичных
Комплементарность цепей составляет химическую основу важнейших функций ДНК — хранения и передачи наследственных признаков. При делении клеток двойная спираль ДНК раскручивается и разделяется на две части. На каждой отдельной цепи, как на матрице, происходит биосинтез новой цепи ДНК с учетом принципа комплементарности. Вновь образовавшаяся цепь не идентична, но комплементарна исходной матрице. В результате воссоздаются две новые двойные спирали ДНК. Такой процесс называется репликацией, и он лежит в основе обеспечения дочерних клеток молекулами ДНК, идентичных
с ДНК родительских клеток. Аналогичным образом на деспирализованном участке цепи ДНК в ядре происходит синтез молекулы матричной (информационной) РНК
(и-РНК), которая затем сама служит матрицей для биосинтеза белка в цитоплазме. Возникающая цепь и-РНК комплементарна той цепи ДНК, на которой она синтезируется. При этом адениновому основанию в ДНК будет соответствовать урациловое основание в РНК, а в качестве углеводного остатка в цепи РНК будет использоваться рибоза. Синтез и-РНК является по существу переписыванием, транскрипцией генетической информации с ДНК на и-РНК. Генетическая информация, т. е. информация о синтезе определенных белков, закодирована в нуклеотидной последовательности ДНК. Одну аминокислоту кодирует трехнуклеотидная последовательность, поэтому код
называют триплетным. Три нуклеотида, контролирующие включение данной аминокислоты в определенный белок в процессе его биосинтеза, называются кодоном.
Третичная структура ДНК. Выделить и изучить нативную молекулу ДНК из ядер эукариотических организмов чрезвычайно трудно, так как молекулы ДНК разрушаются нуклеазами тканей и подвергаются деструкции в условиях выделения.
К настоящему времени удалось выделить в неповрежденном виде ДНК вирусов, митохондрий, некоторых бактерий. Оказалось, что двойная спираль ДНК на некоторых участках может подвергаться дальнейшей спирализации с образованием суперспирали или открытой кольцевой формы.
 |
Суперспирализованная структура обеспечивает экономичную упаковку огромной молекулы ДНК в хромосоме. У человека ДНК клетки организовано в 23 пары хромосом. Средняя протяженность ДНК хромосомы, включающая 130 млн пар оснований, имеет среднюю длину 5 см. Понятно, что уместить такой длины ДНК в ядре можно лишь путем ее определенной упаковки. Многократная спирализация ДНК, сопровождающаяся образованием комплексов с белками, и представляет собой ее третичную структуру. В результате образования такой структуры происходит уменьшение размеров ДНК в 100 тысяч раз. Основным компонентом клеточного ядра является хроматин. Фибриллы хроматина представляют собой структуры, напоминающие бусы на нитке: небольшие, около 10 нм глобулы, связанные друг с другом отрезками ДНК длиной около 20 нм. Эти глобулы получили название нуклеосом. Нуклеосомы состоят из ядра, представленного 8 молекулами гистонов и навитой на него по поверхности молекулы ДНК, делающей 1,75 оборота. Структура нуклеосомы поддерживается гистоном Н1. Количество таких частиц в гаплоидном наборе ДНК человека может составлять величину равную 1,5×107. Одновременно были обнаружены и фибриллы хроматина
с диаметром 25–30 нм, которые могут обратимо превращаться в фибриллы диаметром 10 нм. Полагают, что нить плотно упакованных нуклеосом диаметром 10 нм, образует в свою очередь спиральные витки с шагом спирали около 10 нм. На один виток такой суперспирали приходится 6–7 нуклеосом.
Такие 25–30-нанометровые глобулы получили название нуклеомеров. Нуклеомерный уровень укладки хроматина обеспечивает 40-кратное уплотнение ДНК.
Как нуклеосомный, так и нуклеомерный уровни компактизации ДНК хроматина осуществляются за счет гистоновых белков, которые соединяются неспецифически с ДНК в бороздках. В ДНК встречаются положительные и отрицательные супервитки, образованные
за счет скручивания по часовой или против часовой стрелки двойной спирали. Дальнейшие более высокие уровни организации хроматина, приводящие к еще большей компактизации, связаны со специфическим взаимодействием особых участков ДНК с белками негистоновой природы. Это специфическое связывание приводит к дальнейшему формированию в этих участках больших петель или доменов, на электронных микрофотографиях они выглядят
в виде розеткообразных образований. Эти образования, состоящие из многих петель
30-нанометровых фибрилл, соединяющихся в общем плотном центре, получили название хромомер. Размер отдельной петли совпадает с размером средних репликонов (единиц репликации ДНК) и может соответствовать одному или нескольким генам. На хромосому
в среднем приходится более 2000 таких петельных доменов ДНК. В своих основаниях петли ДНК связаны негистоновыми белками ядерного матрикса. В их состав могут входить как ферменты репликации ДНК, так и транскрипции.
Такая петельно-доменная структура хроматина обеспечивает не только компактизацию хроматина, но и организует функциональные единицы хромосом — репликоны и транскрибируемые гены.
ВИДЫ РИБОНУКЛЕИНОВЫХ КИСЛОТ
Известны несколько видов клеточных РНК: транспортная РНК (т-РНК), информационная РНК (и-РНК), рибосомная РНК (р-РНК), si РНК и другие. Они различаются местоположением в клетке, составом и размерами, а также своими функциями. В составе нуклеопротеинов ядра и цитоплазмы обнаруживаются небольшие, стабильные молекулы РНК. Кроме клеточных РНК, имеются вирусные РНК, входящие в состав многих вирусов животных и растений.
На долю т-РНК приходится 10–20 % от суммы клеточных РНК; их молекулярная масса 30 000, цепь включает 75–90 нуклеотидных звеньев. Основная роль т-РНК состоит в том, что они транспортируют аминокислоты из цитоплазмы к месту синтеза белка — в рибосомы. Число т-РНК превышает число α-аминокислот, участвующих в построении белков.
В класс малых si РНК ( small interfering — малые интерферирующие РНК) включают молекулы, содержащие от 20 до 30 нуклеотидов. Особенностью этих молекул является то, что они, в отличие от большинства других клеточных РНК, состоящих всего из одной цепи нуклеотидов, являются двунитчатыми. Нуклеотиды противоположных цепей si РНК соединяются друг с другом по тем же принципам комплементарности, которые формируют двухцепочечные структуры ДНК в хромосомах. Кроме того, по концам каждой из цепей si РНК всегда остается два неспаренных нуклеотида. В 2002 г. была установлена их способность
подавлять экспрессию генов у животных.
В 2006 г. за исследование механизма действия si РНК, выражающегося в нейтрализации отдельных генов на этапе передачи генетической информации и синтеза белка, двум американским ученым Эндрю Файеру и Крейгу Мелло была присуждена Нобелевская премия в области физиологии и медицины. Это достижение открывает заманчивые перспективы в использовании данной технологии в генной инженерии при изучении роли отдельных генов, путем их «выключения», а также лечении ряда вирусных, эндокринных и других заболеваний.
НАРУШЕНИЯ, ВОЗНИКАЮЩИЕ В ДНК ПОД ВЛИЯНИЕМ
ФАКТОРОВ ОКРУЖАЮЩЕЙ СРЕДЫ
Уже на ранних стадиях эволюции, очевидно, ДНК заменила РНК в качестве носителя генетической информации. Это обусловлено большей устойчивостью ДНК, связанной с заменой рибозы на дезоксирибозу, и двухцепочечным строением ДНК. Благодаря гидрофобным взаимодействиям и водородным связям между двумя комплементарными полинуклеотидными цепями ДНК, реакционноспособные азотистые основания, последовательность
которых кодирует информацию, становятся менее доступными.
Однако, несмотря на свои особенности строения, ДНК постоянно подвергается химическим изменениям, как спонтанным, так и индуцируемым мутагенами и даже клеточными метаболитами. Еще одной причиной повреждения ДНК являются радиация и ультрафиолетовое облучение. Большинство изменений ДНК несовместимо с нормальным функционированием клеток: они либо приводят к вредным мутациям, либо блокируют репликацию ДНК и вызывают гибель клеток. Поэтому все клетки имеют специальные системы репарации ДНК.
В ДНК сравнительно часто и спонтанно происходят апуринизация и дезаминирование оснований. Так, ДНК каждой клетки человеческого организма ежедневно теряет около 5000 пуринов. Результатом апуринизации является АР-сайт (англ. Apurinic-apyrimidinic) — дезоксирибоза, лишенная основания.

При дезаминировании цитозин превращается в урацил, аденин — в гипоксантин,
а гуанин — в ксантин.

 Наиболее существенные нарушения считывания информации происходят при дезаминировании цитозина и аденина: обе реакции после репликации приводят к мутациям. Чаще всего дезаминируется цитозин; в ДНК каждой клетки за день происходит около 100 таких событий. При дезаминировании азотистых оснований, содержащих аминогруппу, образуются основания, не характерные для ДНК. Это обстоятельство позволяет репаративной системе клетки узнавать продукт дезаминирования и удалять его. Очевидно, что именно поэтому в ДНК, в отличие от РНК, вместо урацила присутствует тимин: урацил неотличим от продукта спонтанного дезаминирования цитозина.
Наиболее существенные нарушения считывания информации происходят при дезаминировании цитозина и аденина: обе реакции после репликации приводят к мутациям. Чаще всего дезаминируется цитозин; в ДНК каждой клетки за день происходит около 100 таких событий. При дезаминировании азотистых оснований, содержащих аминогруппу, образуются основания, не характерные для ДНК. Это обстоятельство позволяет репаративной системе клетки узнавать продукт дезаминирования и удалять его. Очевидно, что именно поэтому в ДНК, в отличие от РНК, вместо урацила присутствует тимин: урацил неотличим от продукта спонтанного дезаминирования цитозина.
Многие вещества, обладающие канцерогенным действием, алкилируют, например, метилируют, основания ДНК. Наиболее частые продукты этих реакций — О6-метил-гуанин, 7-метилгуанин и 3-метил-аденин. Первое из этих изменений мутагенно. А два других делают более лабильной N-гликозидную связь и могут способствовать апуринизации.
 При ряде воздействий может происходить размыкание имидазольного цикла пуринового основания. Образующийся при этом продукт — формамидопиримидин создает затруднения для репликации ДНК.
При ряде воздействий может происходить размыкание имидазольного цикла пуринового основания. Образующийся при этом продукт — формамидопиримидин создает затруднения для репликации ДНК.
Основным нарушением, возникающим под действием ультрафиолетового излучения, является насыщение двойных связей азотистых оснований, нарушение ароматичности и плоскостной структуры. В результате образуются пиримидиновые димеры из двух соседних пиримидинов цепи ДНК.
Метаболизм азотистых оснований

Статья будет посвящена базовому, но крайне интересному разделу биохимии — метаболизму азотистых оснований. Сегодня мы рассмотрим то, как из аминокислот и глюкозы получается материал, кодирующий нашу биологическую сущность, — нуклеиновые кислоты (ДНК и РНК). Это понадобится нам для понимания очень важных вещей. Например, как одна мутация в метаболизме нуклеотидов способна подарить ребенку тяжелейший иммунодефицит или сделать из него глубокого инвалида. Почему не живут долго и счастливо многие из тех, кто злоупотребляет вином, красным мясом, шоколадом и прочими радостями. Почему иногда умирают (во всяком случае, часто умирали раньше) при проведении противоопухолевой химиотерапии. В статье, посвященной нарушению метаболизма азотистых оснований, ты узнаешь и о редких заболеваниях, о которых забывают 90 % врачей, принимая их за другие, более известные и очевидные. Но для начала надо постичь базу, ведь без понимания метаболизма азотистых оснований перечисленные мною факты и заболевания будет крайне сложно понять. Приступим.
Немного терминологии
Нуклеотидом мы будем называть соединение, представляющее собой совокупность трех частей [3]:
Уберем фосфат — получим нуклеозид. Уберем еще и рибозу и получим азотистое основание — пуриновое или пиримидиновое «кольцо». Эта терминология необходима для понимания названий целого ряда ферментов, которыми сегодня мы будем развлекать наш мозг.
Попадая в твой ЖКТ, в 12-перстную кишку, нуклеиновые кислоты, содержащиеся в пище, расщепляются рибонуклеазами, нуклеотидазами и прочими ферментами поджелудочной железы. Продукты расщепления в виде нуклеотидов и нуклеозидов (иногда и вовсе азотистых оснований) попадают в энтероциты (клетки слизистой оболочки тонкой кишки).
В энтероцитах есть ксантиноксидаза, которая переводит пурины (аденин, гуанин) в мочевую кислоту. Мочевая кислота, как правило, стремится всосаться в кровь и выйти с мочой через почки. Может также пропустить себя в просвет кишки и выйти с калом (кстати, это бывает и весьма ощутимо, если имеется декомпенсированная патология почек).
Пиримидины утилизируются своим путем [1, 3], о котором мы поговорим позже. Но главное, что ты должен понять: экзогенных источников нуклеотидов для организма нет. Нуклеотиды, попадающие с пищей, не усваиваются, а лишь дают некоторое количество уратов — солей мочевой кислоты. Не перевелись еще люди, которые ведут активную пропаганду «Нет ГМО» и осуществляют на этом нехилую спекуляцию при продаже пищевых продуктов. Пусть в пище будет хоть трижды перекрещенная и модифицированная ДНК, твоя кишка все равно превратит все в кусок химуса и уратов, и ты даже не поймешь, что съел шедевр генной инженерии…
Образование пуринов. «Откуда же тогда взять мономеры для нуклеиновых кислот? Как и откуда берутся нуклеотиды? Как они используются дальше?» — и еще тысяча вопросов, которые добровольно не пришли бы в твою голову, если бы не предстоящий зачет по биохимии. Именно на них мы будем искать ответ.
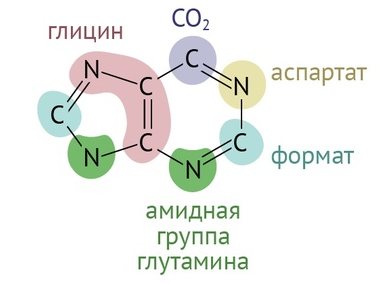
Пурины — гуанин и аденин — формируются буквально из ничего. Все, что нам нужно для создания пуринового кольца — глюкоза (поступающая в пентозо-фосфатный цикл), глицин, две молекулы глутамина, одна молекула аспартата, пара молекул активной метилированной формы фолиевой кислоты + немного терпения и приправы в виде пяти молекул АТФ [1].
Я не хочу грузить тебя серией реакций синтеза пуринов, ее на экзамене полностью никто не спросит, и, скорее всего, для принятия решений в своей работе она тебе тоже не нужна. Но кое-что ты знать должен.
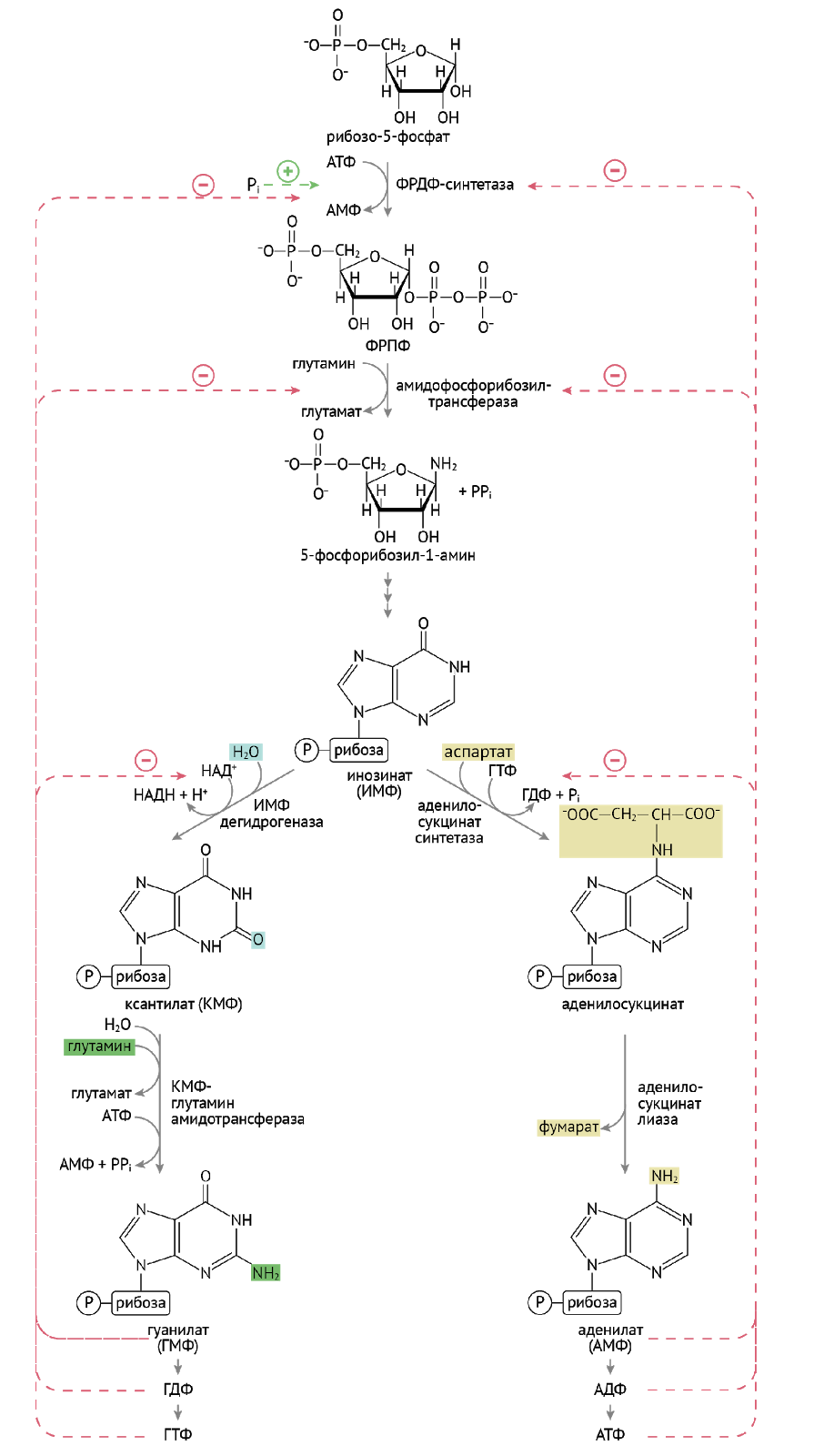
Синтез пуринов проходит два основных этапа: формирование инозинмонофосфата (ИМФ) и получение из ИМФ таких нуклеотидов, как ГМФ и АМФ.
Необходимо знать ключевые моменты этого синтеза.
Первый из них — лимитирующая реакция синтеза пуринов — формирование фосфорибозилпирофосфата (ФРПФ).
(Для тех, кто не понял: лимитирующая реакция — это та реакция, скорость которой определяет скорость всего процесса, а также та, что подвергается наиболее жесткой и сложной регуляции).
Далее следует еще порядка девяти реакций, которые из 5-фосфорибозил-1-амина сделают инозинмонофосфат (ИМФ). На этом закончится первый этап синтеза. Для экзамена и последующей работы тебе нужно знать, какие молекулы участвуют в формировании пуринового кольца, и эти две реакции. Для жизни — и того меньше.
2 этап. Чрезвычайно важен и интересен. Два брата, два параллельно идущих, как ты со своим другом, ̶п̶о̶ ̶н̶а̶к̶л̶о̶н̶н̶о̶й̶ к синтезу АМФ и ГМФ.
Что интересно: конечные продукты этого чрезвычайно интересного (никому) синтеза — ГТФ и АТФ — помогают дружественному параллельному циклу, даруя энергию для синтеза нуклеотида. Несколько ниже приведена иллюстрация вышеописанных биохимических событий.
Распад пуринов и дальнейшая судьба одиноких азотистых оснований
Катаболизм пуринов не менее важная и интересная серия процессов, потому что так же, как и синтез пуринов, их катаболизм является ключевым моментом для понимания патогенеза ряда заболеваний. Большинство нуклеотидов подвергаются повторному использованию (отдать некому, выкинуть жалко), заново включаясь в синтез нуклеотидов. Проследим путь их катаболизма.
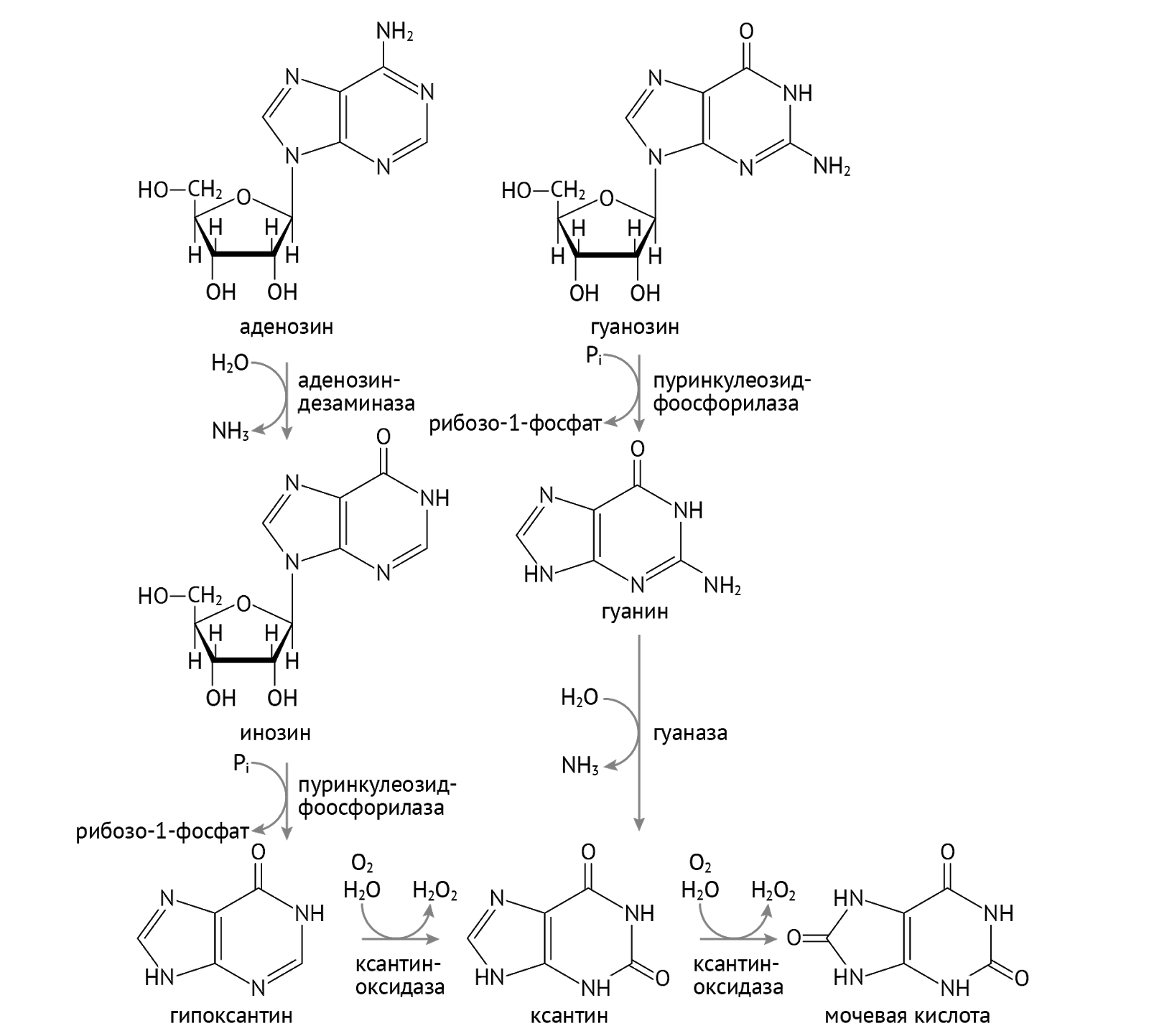
Как мы видим, аденозин (аденин + рибоза) стремится стать гипоксантином. Но на пути к этому ему нужно перевоплотиться назад в инозинмонофосфат. В этом ему поможет аденозиндезаминаза. Хорошо запомни этот фермент, мы к нему не раз еще вернемся. Гипоксантин, кстати, может стать просто ксантином, чем обязан ксантиноксидазе.
Гуанозин считает стадию гипоксантина для себя недостойным (не может смириться с тем, что именно АТФ, а не ГТФ, является основным источником энергии в клетке, старые обиды несчастной молекулы). В силу этого ГТФ не церемонится и становится ксантином, минуя стадию образования гипоксантина. И здесь перед ним встает выбор из двух путей. Путь 1: гипоксантин и ксантин станут мочевой кислотой и оставят после себя лишь воспоминания, удалившись в мочу (или в суставы и почечные канальцы в виде уратов). Путь 2: камбэкнуться в ИМФ (а затем в АМФ) и ГМФ. В этом им поможет братан, которого во дворе прозвали гипоксантингуанинфосфорибозил-трансферазой (ГГФРТ).
Получившиеся вновь ГМФ и ИМФ (🡪 АМФ) превращаются в ГТФ и АТФ путем присоединения фосфорных остатков. Здесь все просто.
Резюме: для нас клиническое значение будут иметь нижеперечисленные ферменты:
Их значение для реализации той или иной патологии будет обсуждаться в соответствующей статье.
Синтез пиримидинов
Здесь все проще и сложнее одновременно. Реакций здесь меньше, они понятнее, а значит, тебя могут спросить их на экзамене. Не пугайся, а просто смотри и запоминай.
С карбомоилфосфатом мы еще встретимся. Забегая далеко вперед (дальше, чем твое отчисление) сообщу тебе, что это соединение – исходный продукт для синтеза мочевины в печени. Той самой мочевины, в которую превращается весь «вредный» азот в организме. Советую отнестись внимательнее к следующим ферментам: оротатфосфорибозил-трасфераза и оротатмонофосфат (ОМФ)-декарбоксилаза. Нарушение их функции — ключевой момент в реализации такого патологического состояния, как оротовая ацидурия.
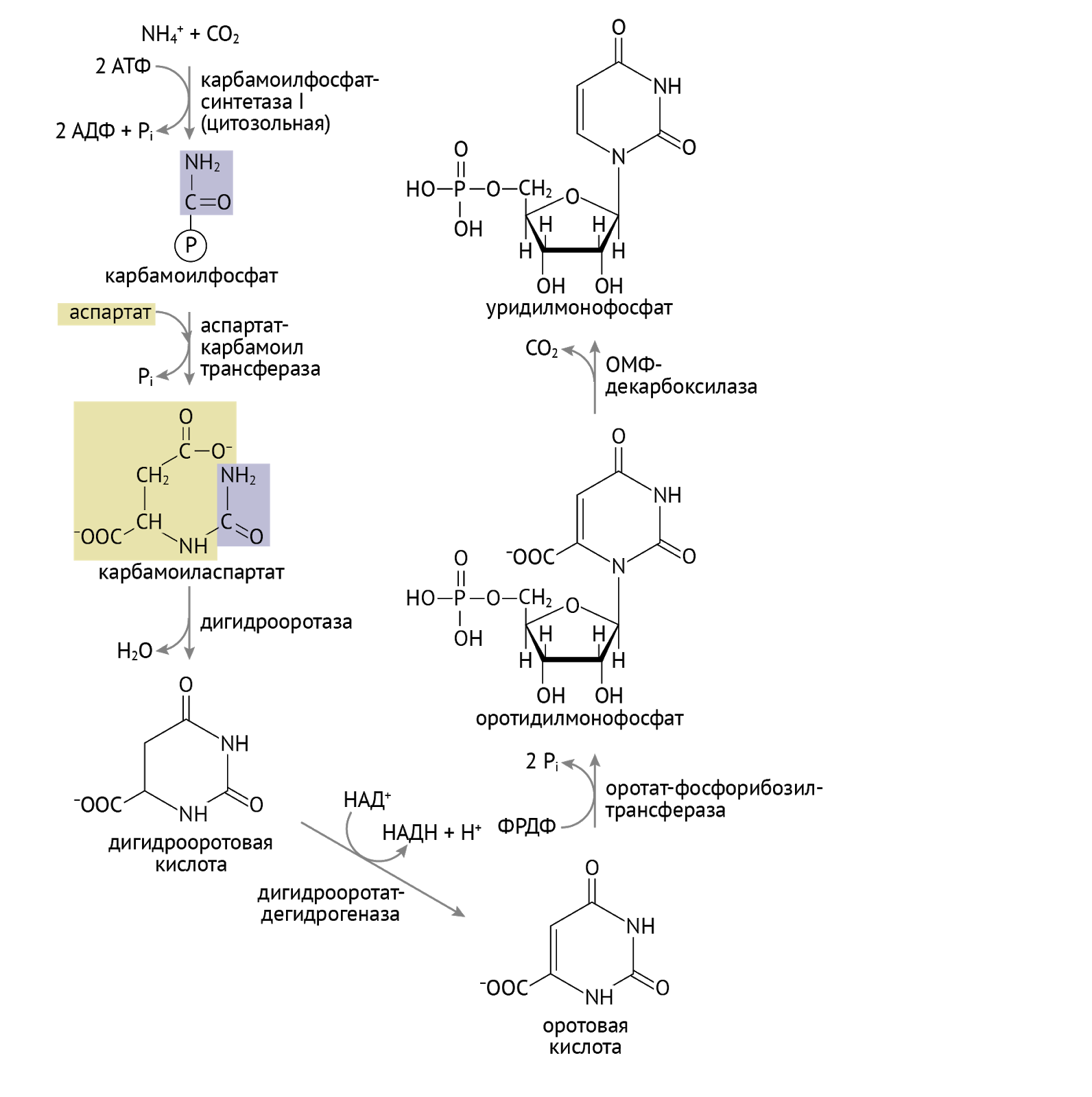
Взглянув на схему, мы видим, что конечным продуктом этого этапа является уридинмонофосфат (УМФ). С обостренным чувством собственной важности УМФ дает начало:
Все образованные соединения — АТФ, ГТФ, ЦТФ, УТФ — используются для синтеза рибонуклеиновых кислот, функцию которых ты должен был помнить не давеча как с третьего класса средней школы. Они имеют важное значение в матричных биосинтезах: в транскрипции ДНК, синтезе белка.
Но у нас есть еще и ДНК. Та штука, которая определяет твой фенотип и все, что из него вытекает. ДНК-полимераза установила фейс-контроль для синтеза ДНК. Он заключается в том, что все субстраты должны быть дезоксирибонуклеотидом [1, 3].
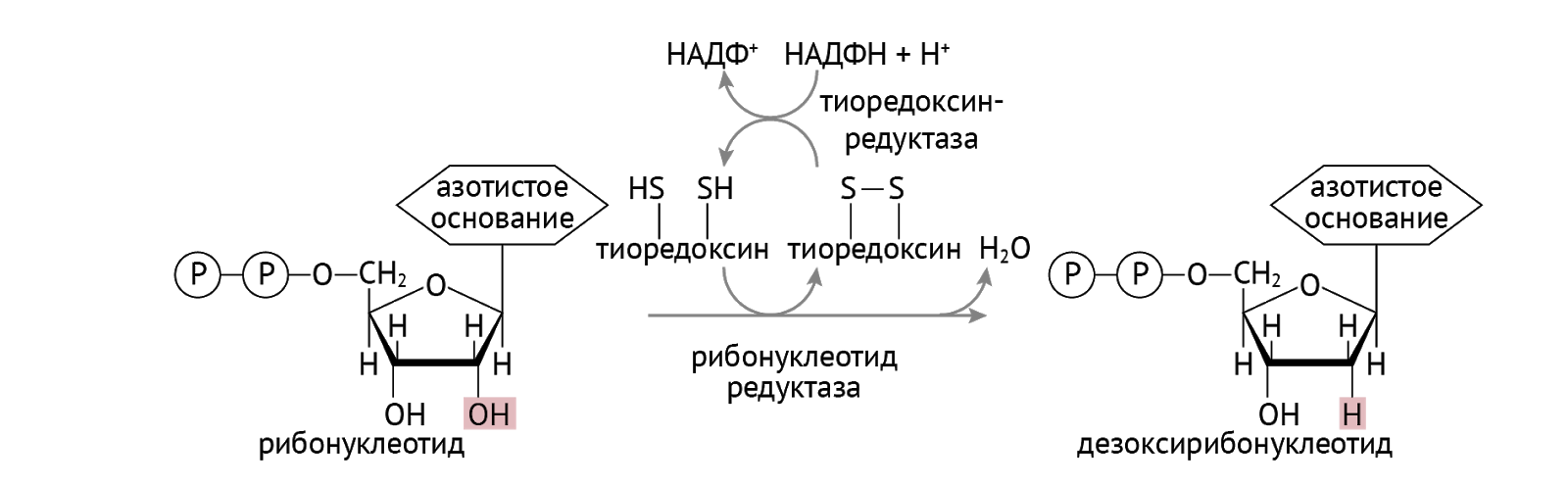
Есть исключение из правил. Это касается УТФ. Чтобы попасть в ДНК, ему нужно притвориться не просто дезоксирибонуклеотидом. Ему нужно притвориться ТТФ (тимидинтрифосфат). Что он и делает:
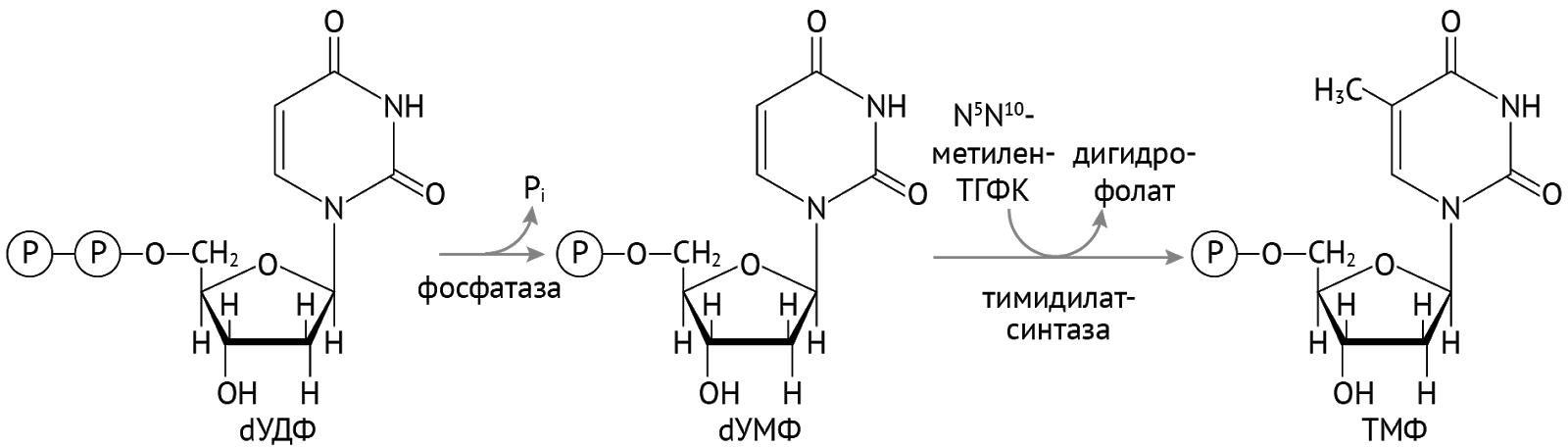
Далее остается присоединить остатки фосфорной кислоты и получить тимидинтрифосфат [1].
Важно отметить, что тимидилатсинтаза и реакция образования ТМФ являются чрезвычайно важными в отношении синтеза ДНК. Также здесь можно заметить, что необходимым кофактором является тетрагидрофолиевая кислота.
Именно данная реакция является мишенью метотрексата — цитостатического противоопухолевого препарата, являющегося антиметаболитом — структурным аналогом фолиевой кислоты, который проявляет к тимидилатсинтазе недвусмысленное сродство. Связываясь с ней, метотрексат останавливает реакцию образования ТМФ, тем самым обрывая синтез ДНК в самом зародыше — на этапе образования ее структурных компонентов. Метотрексат нашел применения в онкологии (преимущественно при гемобластозах) и в ревматологии (ревматоидный артрит [5]).
Распад пиримидинов
В нашей жизни ничего не вечно. Пиримидины тоже гибнут, но распадаясь, оставляют после себя ряд предшественников, которые участвуют в метаболизме, но уже другими путями.
Комментируя сложную схему распада пиримидинов, я акцентирую внимание на двух важных вещах:
Биологическое значение нуклеотидов в клетках. Функции, выходящие за рамки синтеза нуклеиновых кислот
Обсудив с тобой предметы сложной и высокой материи, мы переходим к самому интересному и «вкусному» (только не блевани) разделу настоящей статьи.
Нуклеотиды являются кирпичами для нуклеиновых кислот, что важно для реализации генетической информации и превращения ее в реальный фенотип со своими уникальными морфологическими особенностями. Этот процесс заслуживает отдельной лекции (спойлер — она следующая).
Но этим не исчерпывается значение нуклеотидов.
Детоксикация. Кое-кто из них используется в организме для детоксикации ксенобиотиков — лекарств, токсинов и иных химикатов, поступающих в организм, а также для детоксикации липофильных (т. е. растворимых в липидах и нерастворимых в воде) соединений: гормонов, билирубина. Суть детоксикации (обезвреживания) — перевести плохо растворимое в воде вещество в хорошо растворимое, что необходимо для последующего выведения через почки с мочой из организма. Достигается это присоединением полярной химической группы к исходному хулигану. Сначала формируется гидроксильная группа (если таковой нет, это этап образования более полярных соединений). А затем наступает этап конъюгации… Посмотрим, как это выходит на примере тетрагидрокортизола (рис. 11) [2].
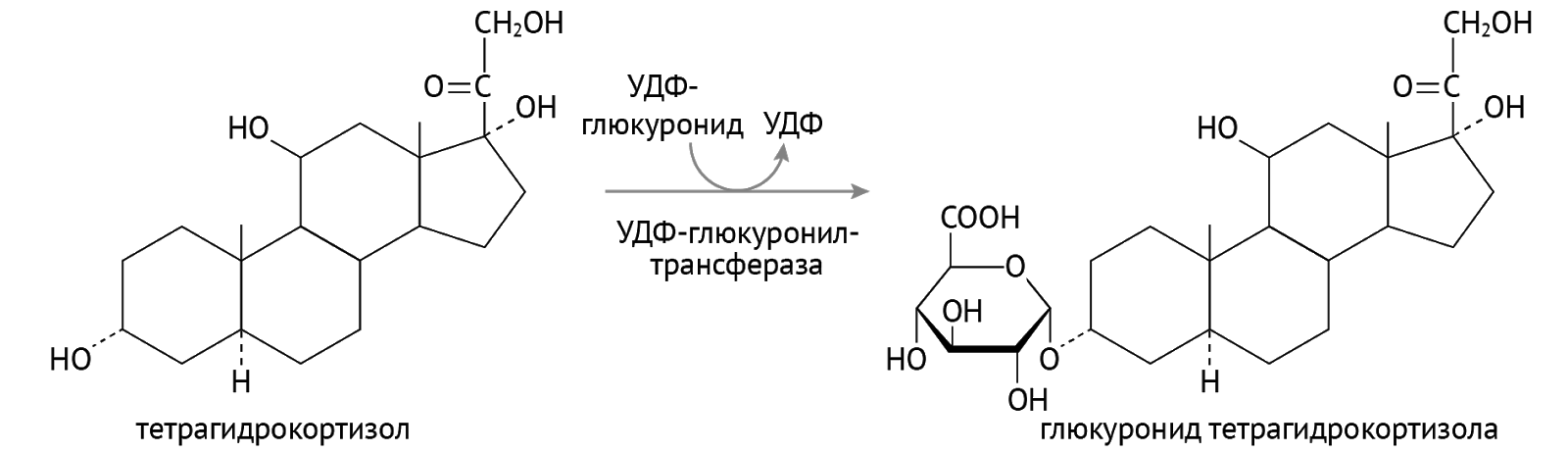
А вот схема для менее дотошных, чем автор данной лекции.
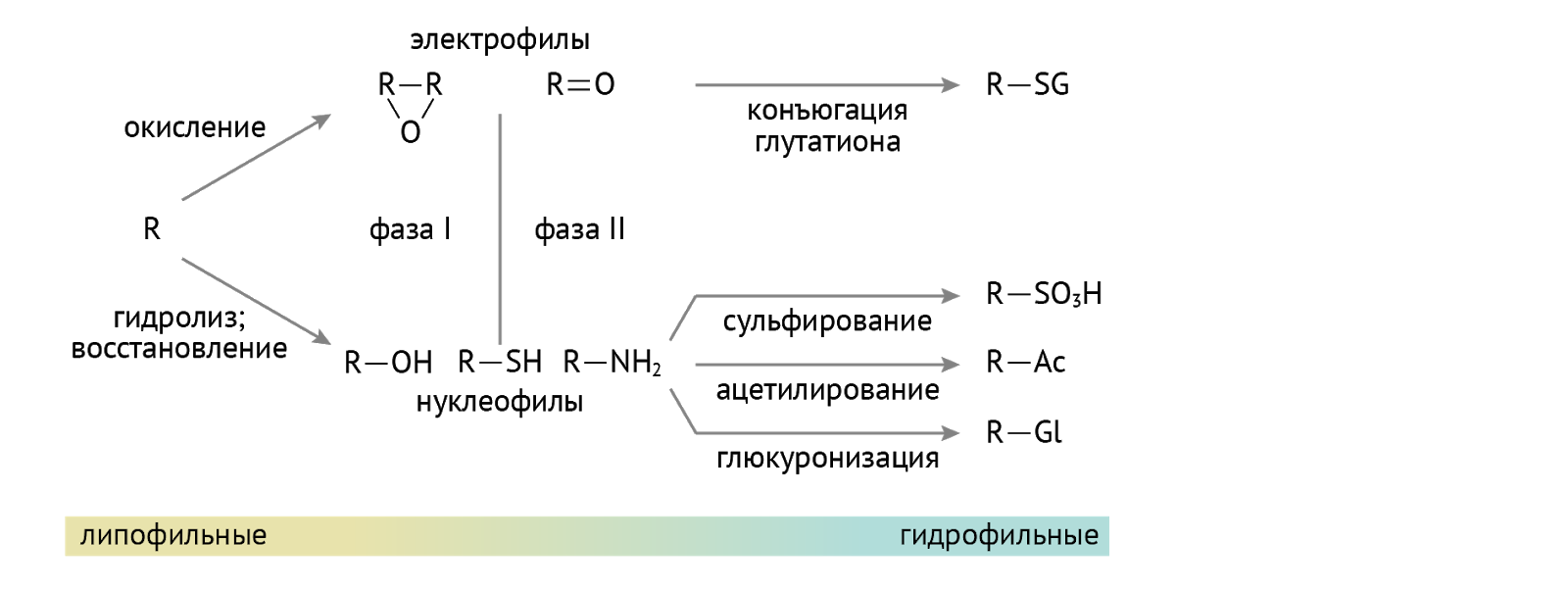
Никто: «Ух ты, классно и понятно».
Твой экзаменатор: «А откуда ты возьмешь такую форму нуклеотида, которая будет способна конъюгировать с ксенобиотиками?».
Ты:
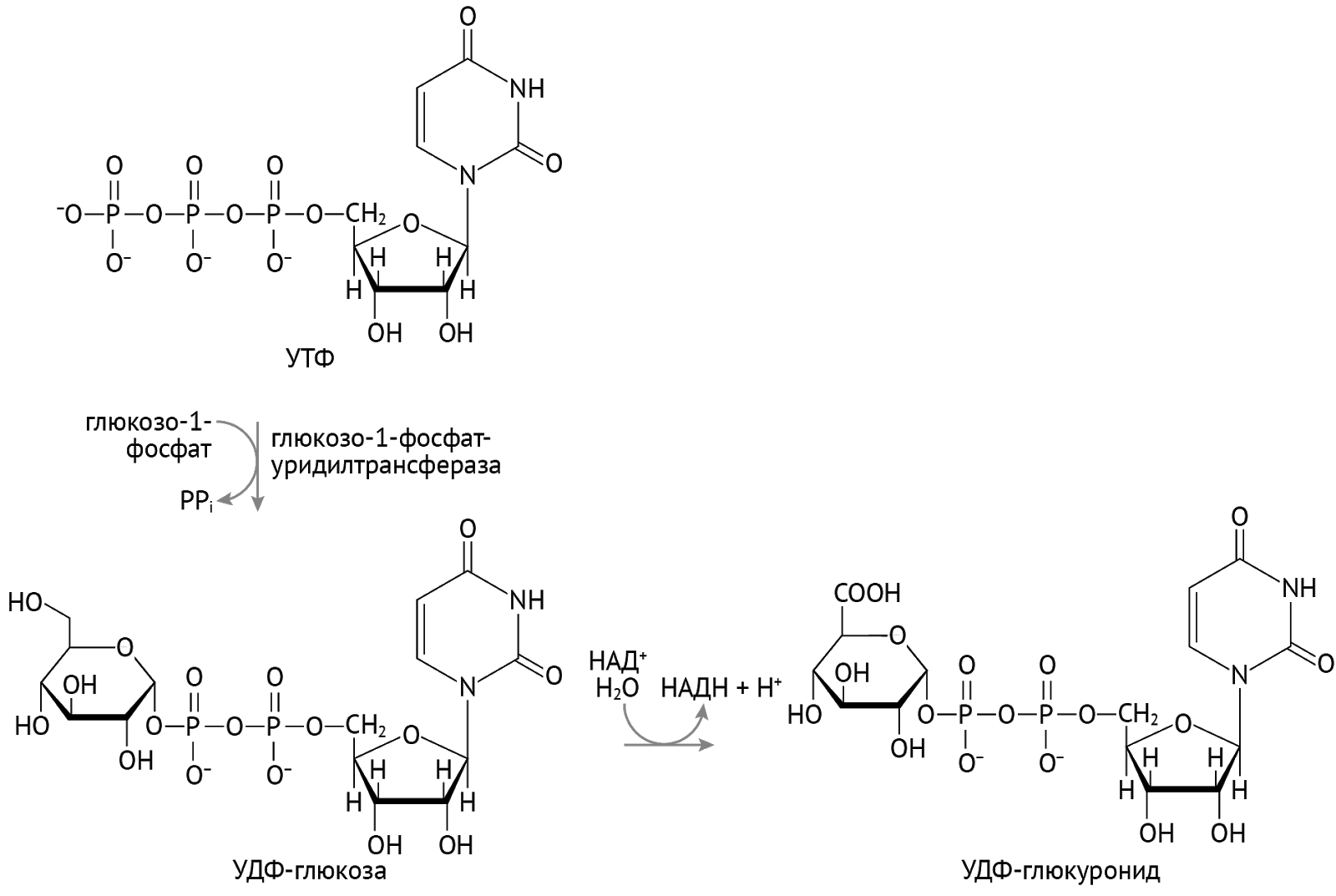
Важное значение эта форма УДФ имеет и для метаболизма билирубина: благодаря УДФ-диглюкурониду токсичный липофильный билирубин становится полярным, растворимым в крови и моче, конъюгированным билирубином [1, 2].
Гликоген. Неоценимое значение УДФ имеет и для образования гликогена.
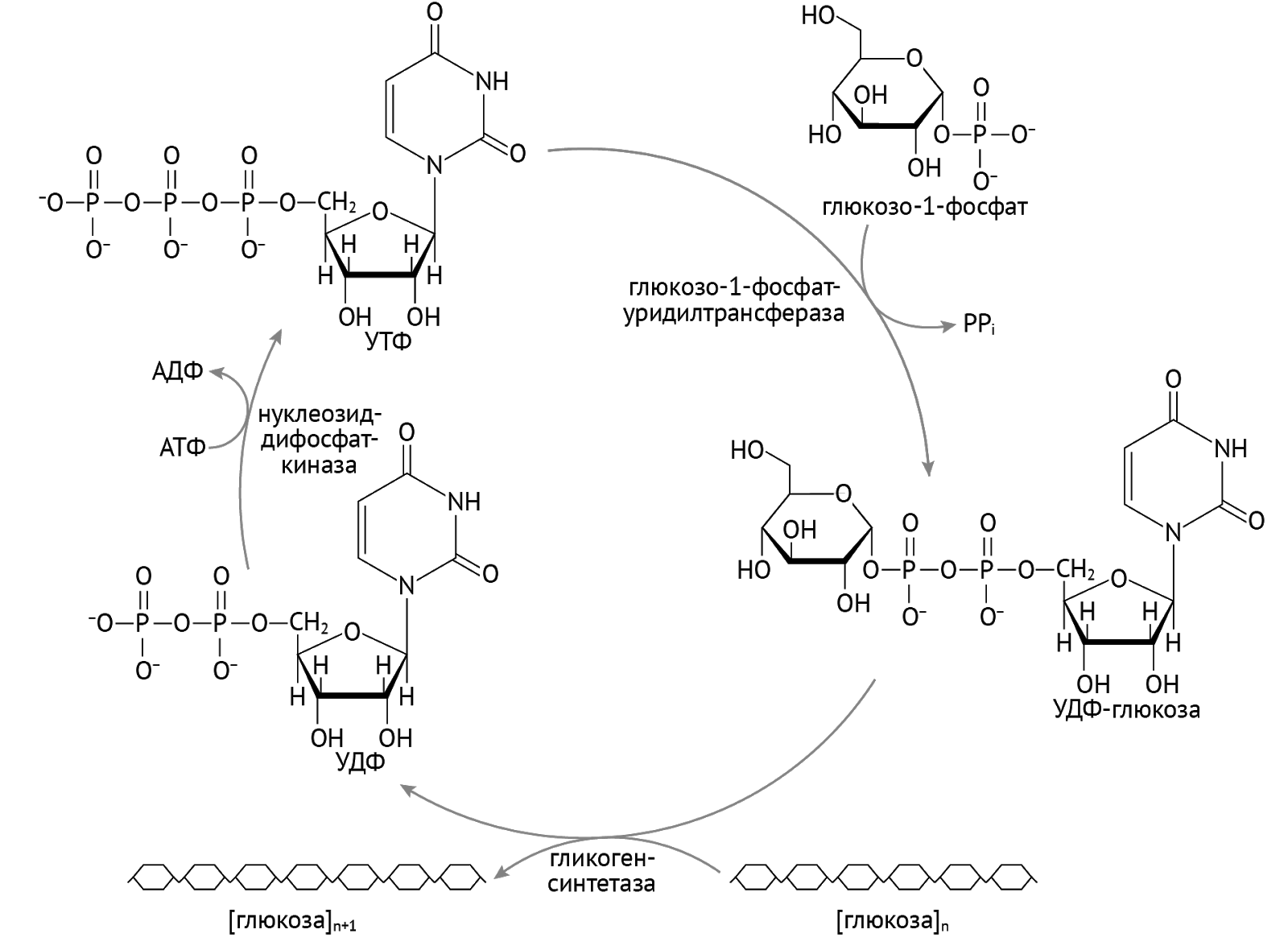
Гликоген — это сравнительно инертная макромолекула, которая является резервуаром источника энергии в печени и мышцах. Через восприятия сигнала от инсулина глюкоза посредством активации (в виде присоединения фосфорного остатка и изомеризации) прыгает на гликоген благодаря посредничеству УТФ [1].
Вторичные сигнальные молекулы. Нуклеотиды, в частности, цАМФ и цГМФ, играют важную роль в процессе реализации клеточных функций и межклеточного взаимодействия. Клетка, будучи кирпичом биологического коммунизма, воспринимает сигналы от вышестоящих регуляторных систем, которые контролируют ее деятельность.
Это может выражаться в:
Такие молекулы, как цАМФ и цГМФ, являются одними из важных вторичных мессенджеров. Клетка, воспринимая сигнал извне (будь то гормон, нейромедиатор или любой другой разбойник), изменяет внутри себя концентрацию этих мессенджеров. Те вмешиваются в серию внутриклеточных последовательных реакций и тем самым усиливают первоначальный сигнал [4].
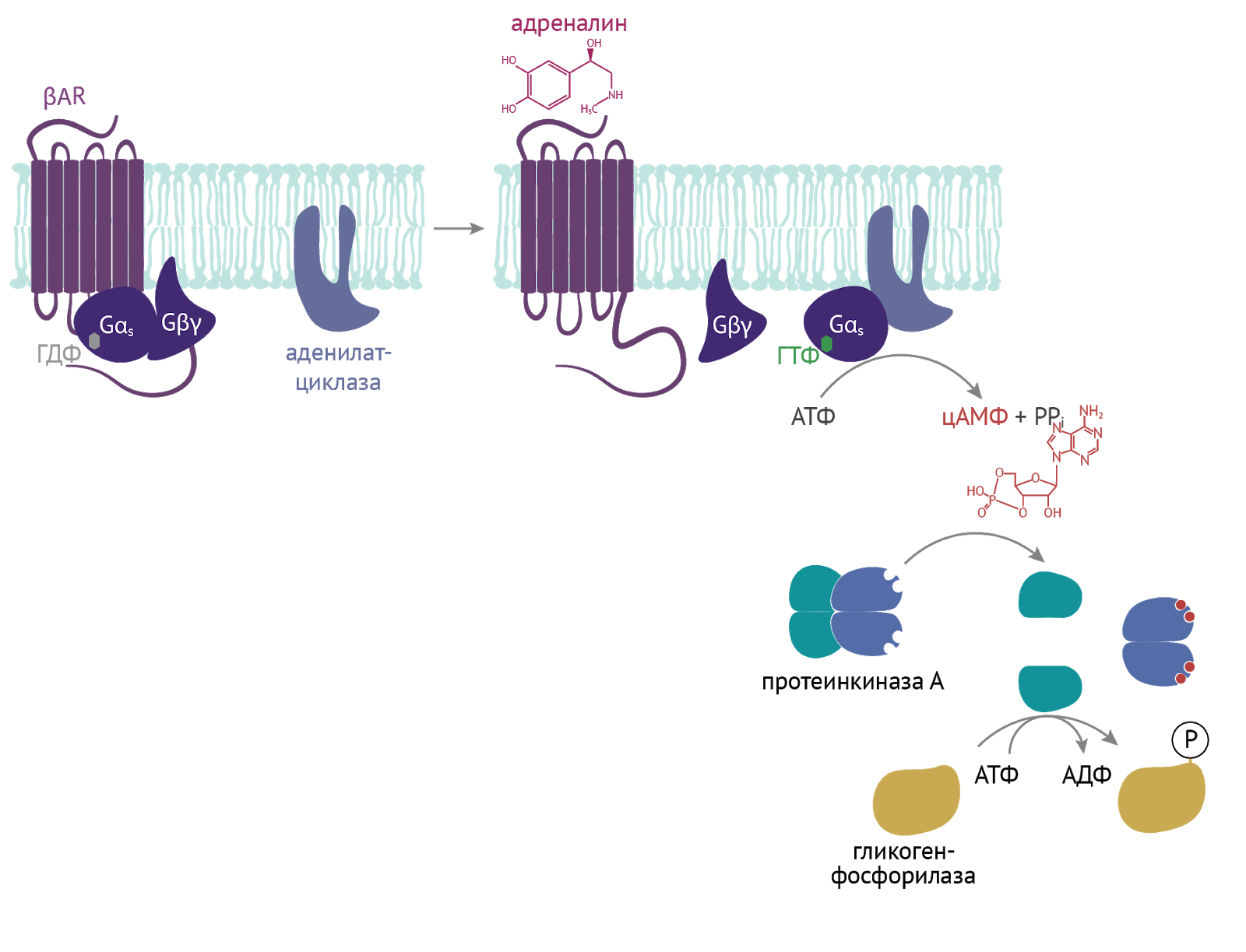
Здесь пример воздействия адреналина на метаболизм гликогена. Концентрация цАМФ растет, что приводит к активации протеинкиназы А. Та активирует гликогенфосфорилазу, что приводит к высвобождению из гликогена глюкозы и получению дополнительного источника энергии для выполнения различных нужд организма [4].
Примеров, когда нуклеотиды играют важную роль в организме, чрезвычайно много. Вспомнить хотя бы о том, кто в клетках является главным источником энергии (АТФ). Нуклеотиды в ходе своего метаболизма могут становиться источником иных метаболитов, которые могут принимать участие в энергетическом метаболизме (α-кетоглутарат, пировиноградная кислота 🡪 ацетил-Коа) и так до бесконечности. Придет время, и мы обсудим те случаи, когда метаболизм пуринов и пиримидинов в силу тех или иных причин нарушается [1].
Иногда это имеет очень серьезные последствия…