Фотосинтез
Типы питания

Фотосинтез

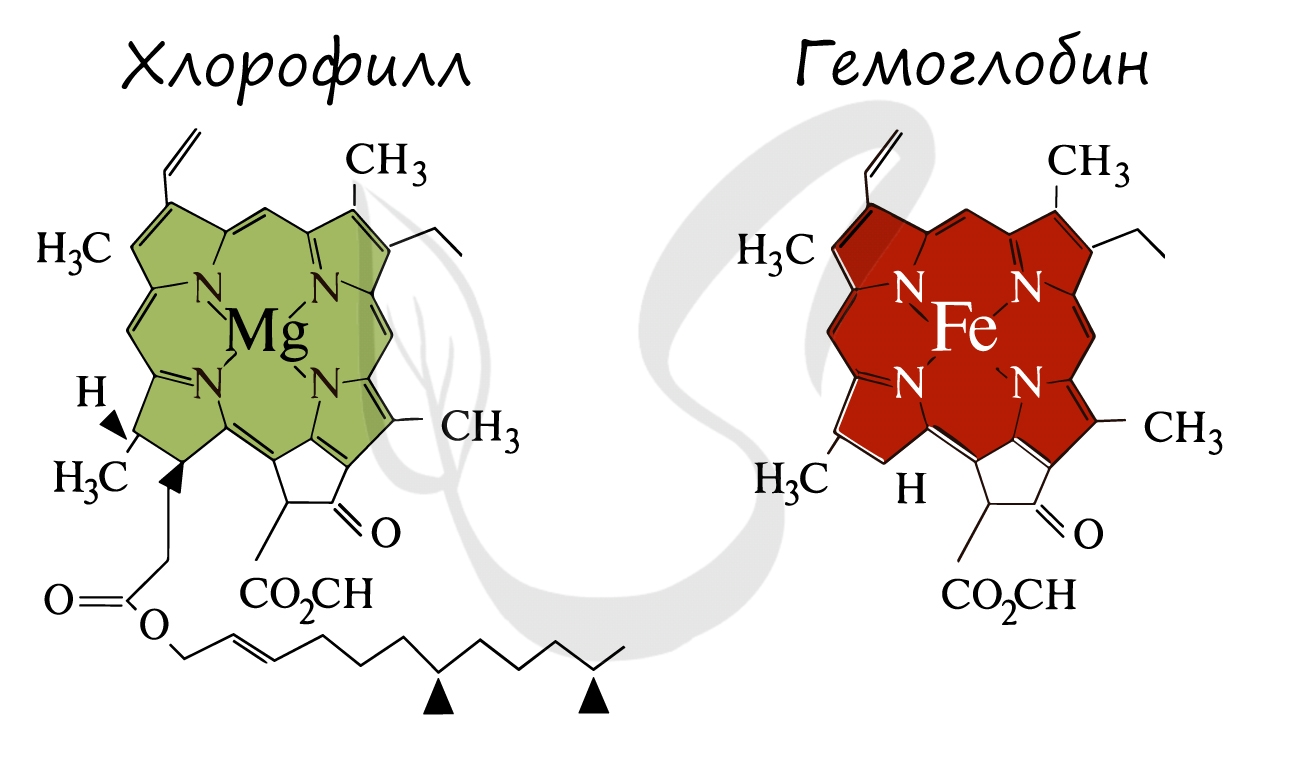
Ниже вы увидите сравнение строения хлорофилла и гемоглобина. Обратите внимание, что в центре молекулы хлорофилла находится ион Mg.
В высшей степени гениально значение процесса фотосинтеза подчеркнул русский ученый К.А. Тимирязев: «Все органические вещества, как бы они ни были разнообразны, где бы они ни встречались, в растении ли, в животном или человеке, прошли через лист, произошли от веществ, выработанных листом. Вне листа или, вернее, вне хлорофиллового зерна в природе не существует лаборатории, где бы выделялось органическое вещество. Во всех других органах и организмах оно превращается, преобразуется, только здесь оно образуется вновь из вещества неорганического»
Более подробно мы обсудим значение фотосинтеза в завершение этой статьи. Фотосинтез состоит из двух фаз: светозависимой (световой) и светонезависимой (темновой). Я рекомендую использовать названия светозависимая и светонезависимая, так как они способствуют более глубокому (и правильному!) пониманию фотосинтеза.
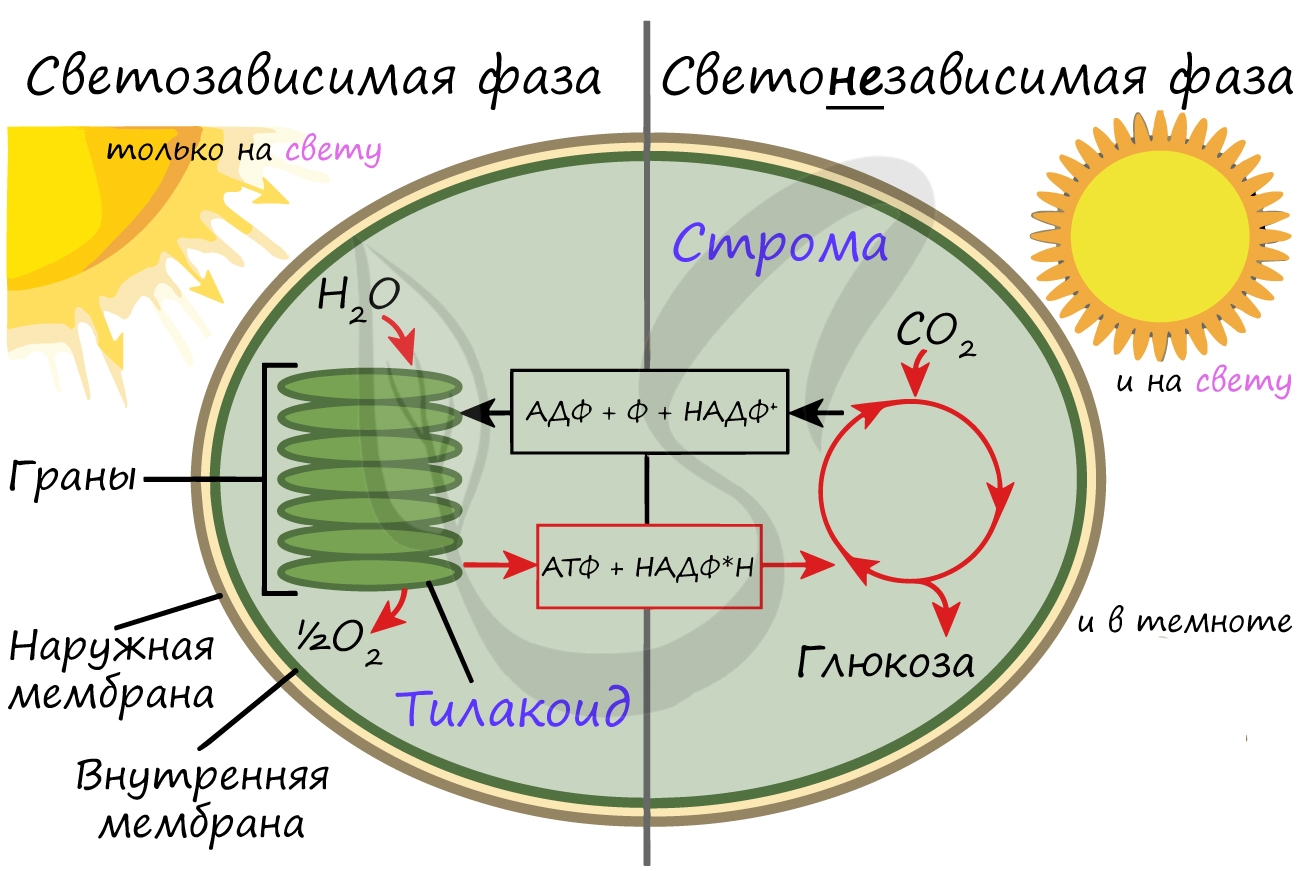
Светозависимая фаза (световая)
Эта фаза происходит только на свету на мембранах тилакоидов в хлоропластах. В ней принимают участие различные ферменты, белки-переносчики, молекулы АТФ-синтетазы и зеленый пигмент хлорофилл.
Хлорофилл выполняет две функции: поглощения и передачи энергии. При воздействии кванта света хлорофилл теряет электрон, переходя в возбужденное состояние. С помощью переносчиков электроны скапливаются с наружной поверхности мембраны тилакоидов, тем временем внутри тилакоида происходит фотолиз воды (разложение под действием света):
Гидроксид-ионы отдают лишний электрон, превращаясь в реакционно способные радикалы OH, которые собираются вместе и образуют молекулу воды и свободный кислород (это побочный продукт, который в дальнейшем удаляется в ходе газообмена).
При достижении критической разницы, часть протонов проталкивается на внешнюю сторону мембраны через канал АТФ-синтетазы. В результате этого выделяется энергия, которая может быть использована для фосфорилирования молекул АДФ:
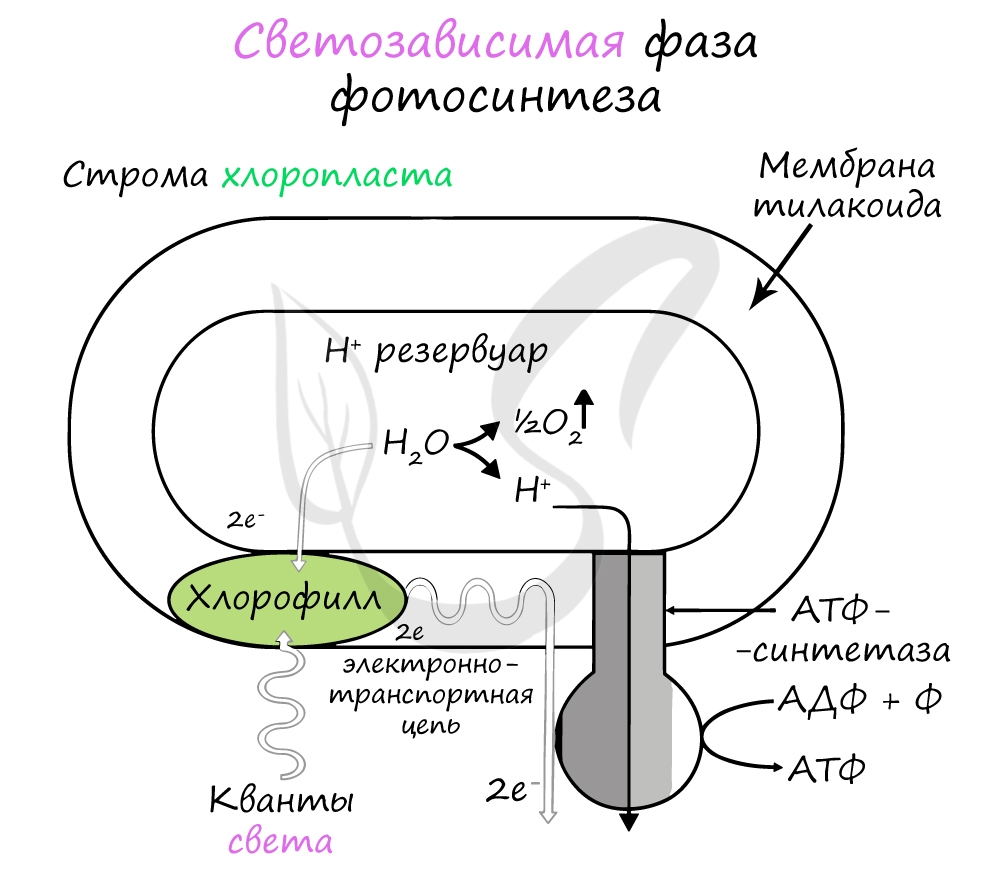
Кислород удаляется из клетки как побочный продукт фотосинтеза, он совершенно не нужен растению. АТФ и НАДФ∗H2 в дальнейшем оказываются более полезны: они транспортируются в строму хлоропласта и принимают участие в светонезависимой фазе фотосинтеза.
Светонезависимая (темновая) фаза
При участии АТФ и НАДФ∗H2 происходит восстановление CO2 до глюкозы C6H12O6. В светонезависимой фазе происходит цикл Кальвина, в ходе которого и образуется глюкоза. Для образования одной молекулы глюкозы требуется 6 молекул CO2, 12 НАДФ∗H2 и 18 АТФ.
Таким образом, в результате темновой (светонезависимой) фазы фотосинтеза образуется глюкоза, которая в дальнейшем может быть преобразована в крахмал, служащий для запасания питательных веществ у растений.
Значение фотосинтеза
Значение фотосинтеза невозможно переоценить. Уверенно утверждаю: именно благодаря этому процессу жизнь на Земле приобрела такие чудесные и изумительные формы, какие мы видим вокруг себя: удивительные растения, прекрасные цветы и самые разнообразные животные.

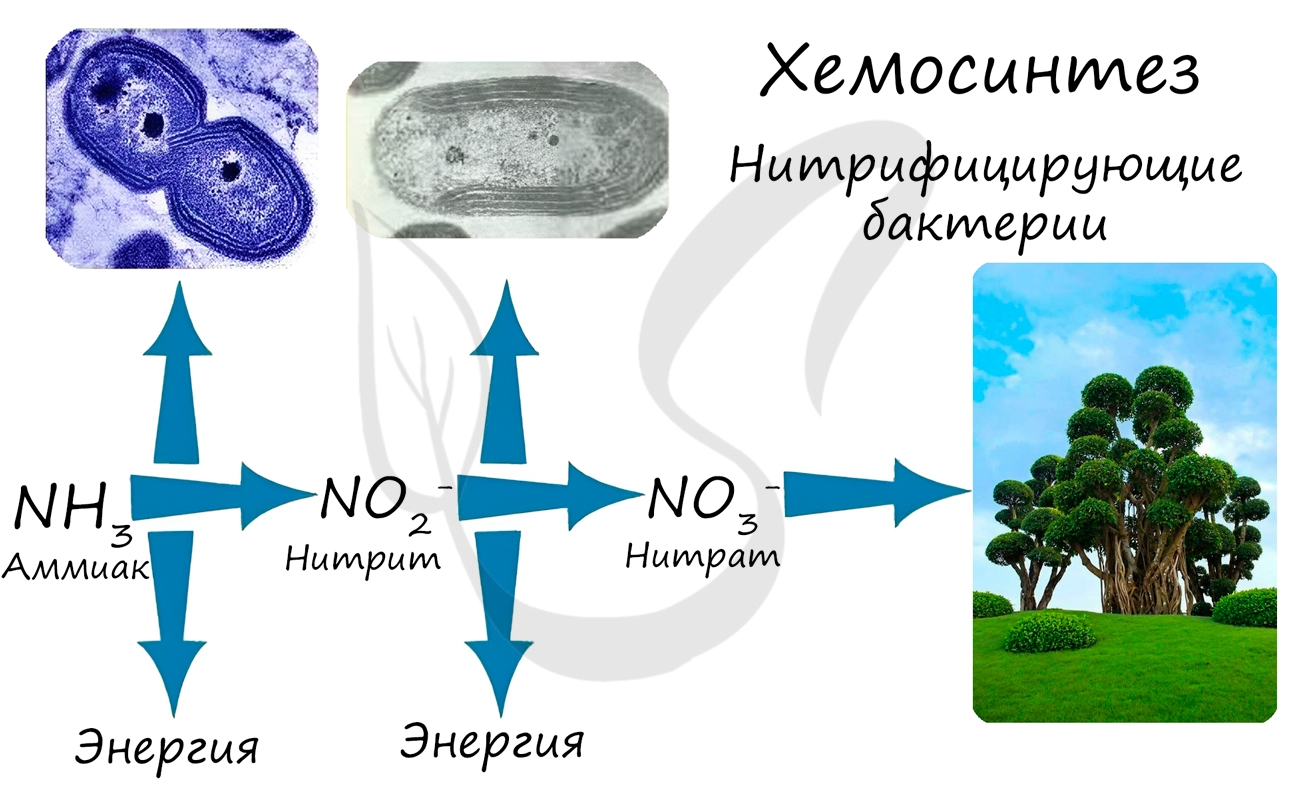
Хемосинтез был открыт русским микробиологом С.Н. Виноградским в 1888 году. Большинство хемосинтезирующих бактерий относится к аэробам, для жизни им необходим кислород.
Значение хемосинтеза
Хемосинтезирующие бактерии являются неотъемлемым звеном круговорота в природе таких элементов как: азот, сера, железо.
Усвоение нитратов происходит за счет клубеньковых бактерий на корнях бобовых растений, однако важно помнить, что клубеньковые (азотфиксирующие) бактерии, в отличие от нитрифицирующих бактерий, питаются гетеротрофно.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
На что затрачивается энергия в процессе фотосинтеза


Фотосинтезом называют процесс преобразования энергии света в энергию химически связей органических соединений с участием хлорофилла. В результате фотосинтеза образуется около 150 млрд тонн органического вещества и приблизительно 200 млрд тонн кислорода ежегодно. Этот процесс обеспечивает круговорот углерода в биосфере, не давая накапливаться углекислому газу и препятствуя тем самым возникновению парникового эффекта и перегреву Земли. Образующиеся в результате фотосинтеза органические вещества частично расходуются гетеротрофными организмами, но значительная их часть в течение миллионов лет образовала залежи полезных ископаемых (каменного и бурого угля, нефти).

К. А. Тимирязев (1843–1920) назвал роль фотосинтеза «космической», поскольку он связывает Землю с Солнцем (космосом), обеспечивая приток энергии на планету.
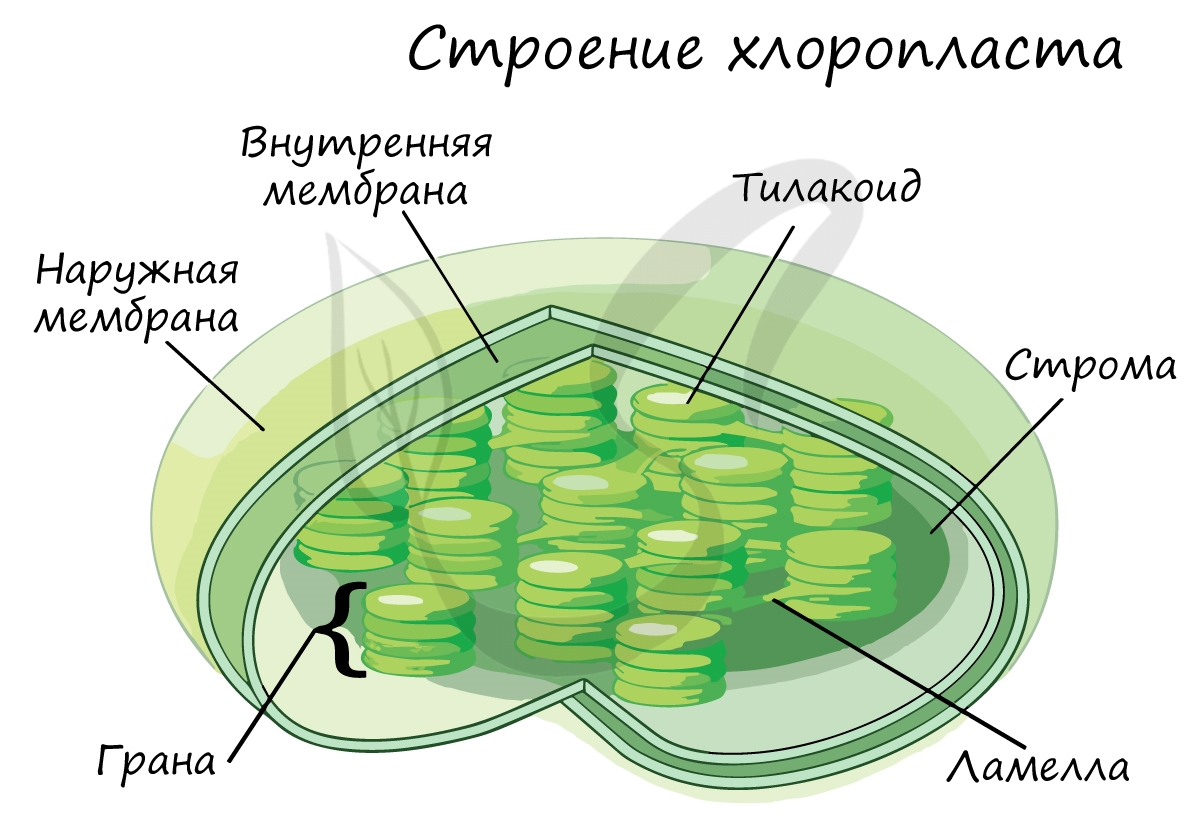
Структурной и функциональной единицей хлоропластов являются тилакоиды – плоские мембранные мешочки, уложенные в стопки (граны) (рис. 3).

Рис. 3. Строение хлоропласта
Отдельные граны соединены друг с другом ламеллами.
В мембранах тилакоидов расположены особые комплексы, в которые входит молекула хлорофилла, а также молекула переносчиков электронов – цитохромов. Мембранная система – это то место, где протекают световые реакции фотосинтеза.
Строма хлоропластов по своему строению напоминает гель – здесь протекают темновые реакции.
Избыток углеводов, образовавшихся в процессе фотосинтеза, запасается в виде зерен крахмала.


Фотосинтез происходит в две фазы, а именно в световую фазу и темновую фазу.
Во время световой фазы происходит образование энергии, которая затем расходуется на темновые реакции. Процесс световой фазы фотосинтеза включает в себя нециклическое фотофосфорилирование и фотолиз воды. В качестве побочного продукта реакции в результате фотолиза воды выделяется кислород. Реакция происходит на мембранах тилакоидов.
Квант красного света, поглощенный хлорофиллом П680 (фотосистема ІІ), переводит электрон в возбужденное состояние (рис. 6). Возбужденный светом электрон приобретает большой запас энергии, вследствие чего перемещается на более высокий энергетический уровень. Такой электрон захватывается акцептором электронов Х, перемещаясь с одной ступени на другую, то есть от одного акцептора к другому, он теряет энергию, которая используется для синтеза АТФ.

Рис. 6. Схема процессов световой фазы фотосинтеза
Место вышедших электронов молекулы хлорофилла П680, занимают электроны воды, так как вода под действием света подвергается фотолизу, где в качестве побочного продукта образуется кислород. Фотолиз происходит в полости тилакоида (рис. 7).

Рис. 7. Фотолиз воды
В фотосистеме І возбужденные электроны под действием фотона света также переходят на более высокий уровень и захватываются акцептором Y. В конце концов, электроны доходят от Y до переносчика – НАДФ, и, взаимодействуя с ионами водорода, выделенными при фотолизе воды, образуют восстановленный НАДФН. НАДФ расшифровывается как – никотинамидадениндинуклеотидфосфат.

Рис. 8. Взаимодействие фотосистемы I и фотосистемы II
Место вышедших электронов в молекуле П700 занимают электроны, полученные от фотосистемы II П680 (рис. 8). Таким образом, на свету электроны перемещаются от воды к фотосистемам II и I, и затем к НАДФ. Такой однонаправленный поток электронов носит название нециклического потока электронов, а образование АТФ, которое при этом происходит, носит название нециклического фотофосфорилирования. Таким образом, в световой фазе образуются АТФ и восстановленный НАДФ, богатые энергией, и в качестве побочного продукта реакции выделяется кислород.
Темновая фаза фотосинтеза. Если световая фаза протекает только на свету, то темновая фаза не зависит от света. Темновая фаза протекает в строме хлоропластов, куда переносятся богатые энергией соединения, а именно АТФ и восстановленный НАДФ, кроме этого, туда же поступает углекислый газ в качестве источника углеводов, который берется из воздуха и поступает в растения через устьица. В реакциях темновой фазы углекислый газ восстанавливается до глюкозы с помощью энергии, запасенной молекулами АТФ и НАДФ.
Превращение углекислого газа в глюкозу в ходе темновой фазы фотосинтеза получило название цикла Кальвина – по имени его первооткрывателя.
Первая стадия фотосинтеза – световая – происходит на мембранах хлоропласта в тилакоидах.
Вторая стадия фотосинтеза – темновая – протекает внутри хлоропласта, в строме.
Суммарное уравнение фотосинтеза выглядит следующим образом. При взаимодействии 6 молекул углекислого газа и 6 молекул воды образуется одна молекула глюкозы и выделяется шесть молекул кислорода. Этот процесс протекает на свету в хлоропластах у высших растений.

Таким образом, фотосинтез – процесс превращения вещества и энергии.
ЭНЕРГЕТИКА ФОТОСИНТЕЗА
1.Энергетика фотосинтеза. Значение отдельных участков солнечного спектра для фотосинтеза
2.1 Фотофизический этап фотосинтеза
2.2 Фотохимический этап
2.3 Циклический и нециклический поток электронов. Фотосинтетическое фосфорилирование
1.Энергетика фотосинтеза. Значение отдельных участков солнечного спектра для фотосинтеза
Важнейшей особенностью процесса фотосинтеза является то, что он протекает с использованием энергии солнечного света.
Лучистая энергия — это энергия электромагнитных колебаний, которая характеризуется определенной длиной волны, частотой колебания и скоростью распространения. Эти величины связаны между собой следующим образом: 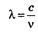 , где λ — длина волны, с — скорость света, величина постоянная, равная 3-10 10 см/с, v — частота колебаний. Длина волны измеряется в нанометрах, а частота колебаний—в герцах (длина волны — расстояние между двумя последовательными пиками одного цикла, частота колебаний — частота появления пиков). Собственно свет, или область электромагнитных колебаний, воспринимаемая человеческим глазом, лежит в пределах длины волн от 400 до 700 нм. Более короткие волны характерны для ультрафиолетовых лучей, а более длинные — для инфракрасных. Известно, что при пропускании луча света через призму он разлагается на отдельные участки, имеющие различную окраску (спектр). В пределах видимого света в зависимости от длины волны окраска лучей различна. Обладая волновыми характеристиками, свет проявляет и корпускулярные свойства. Лучистая энергия излучается и распространяется в виде отдельных дискретных единиц — квантов или фотонов. Квант света обладает энергией, которая прямо пропорциональна частоте колебаний и обратно пропорциональна длине волны
, где λ — длина волны, с — скорость света, величина постоянная, равная 3-10 10 см/с, v — частота колебаний. Длина волны измеряется в нанометрах, а частота колебаний—в герцах (длина волны — расстояние между двумя последовательными пиками одного цикла, частота колебаний — частота появления пиков). Собственно свет, или область электромагнитных колебаний, воспринимаемая человеческим глазом, лежит в пределах длины волн от 400 до 700 нм. Более короткие волны характерны для ультрафиолетовых лучей, а более длинные — для инфракрасных. Известно, что при пропускании луча света через призму он разлагается на отдельные участки, имеющие различную окраску (спектр). В пределах видимого света в зависимости от длины волны окраска лучей различна. Обладая волновыми характеристиками, свет проявляет и корпускулярные свойства. Лучистая энергия излучается и распространяется в виде отдельных дискретных единиц — квантов или фотонов. Квант света обладает энергией, которая прямо пропорциональна частоте колебаний и обратно пропорциональна длине волны 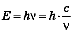 , где h — постоянная Планка. Из этой формулы ясно видно, что величина энергии квантов для разных участков спектра будет различна: чем короче длина волны, тем больше энергия кванта. Энергия квантов разных областей спектра может быть вычислена в электронвольтах или в килоджоулях на моль. 1 моль квантов, или 1 Эйнштейн, соответствует числу квантов, равному 6,02310 23 (число Авогадро). Соответствующие расчеты приведены в таблице 1.
, где h — постоянная Планка. Из этой формулы ясно видно, что величина энергии квантов для разных участков спектра будет различна: чем короче длина волны, тем больше энергия кванта. Энергия квантов разных областей спектра может быть вычислена в электронвольтах или в килоджоулях на моль. 1 моль квантов, или 1 Эйнштейн, соответствует числу квантов, равному 6,02310 23 (число Авогадро). Соответствующие расчеты приведены в таблице 1.
Согласно первому закону фотохимии, только поглощенные лучи могут быть использованы в химических реакциях. В том случае если реагирующие молекулы бесцветны и не поглощают свет, фотохимические реакции могут идти только в присутствии специальных веществ — фотосенсибилизаторов. Фотосенсибилизаторы — вещества, поглощающие энергию света и передающие ее на ту или иную бесцветную молекулу.
Таблица 1 Характеристика отдельных участков спектра
| Цвет | Длина волны, нм | Энергия квантов, кДж/моль |
| Ультрафиолетовый | 471,4 | |
| Фиолетовый | 400-424 | 292,0 |
| Синий | 424-491 | 260,6 |
| Зеленый | 491-550 | 230,5 |
| Желтый | 550-585 | 206,6 |
| Оранжевый | 585-647 | 193,6 |
| Красный | 647-740 | 176,4 |
| Инфракрасный | 85,5 |
Положение о том, что в процессе фотосинтеза могут быть использованы только поглощенные лучи солнечного света, впервые получило экспериментальное подтверждение в опытах К.А. Тимирязева. До этого господствовало ошибочное представление, что наибольшее значение в процессе фотосинтеза имеют желтые лучи солнечного спектра, которые хлорофиллом не поглощаются. К.А. Тимирязев показал, что процесс усвоения СО2 на свету представляет собой фотохимический процесс и подчиняется законам фотохимии. В процессе фотосинтеза вместо связей, обладающих малым запасом энергии, таких, как О—Н, С—О, создаются связи С—С, благодаря этому свободная энергия системы повышается. Эта энергия представляет собой трансформированную солнечную энергию,
К.А. Тимирязевым был разработан точный метод учета процесса фотосинтеза, основанный на одновременном определении объема поглощенного углекислого газа и выделенного кислорода в замкнутом сосуде (эвдиометре). Высокая чувствительность данного метода позволила определить фотосинтез в отдельных участках спектра, в которых благодаря малой их интенсивности фотосинтез шел очень слабо. Опыты К.А. Тимирязева ясно показали, что процесс фотосинтеза проходит именно в тех лучах, которые поглощаются хлорофиллом. Хлорофилл является оптическим сенсибилизатором, поглощающим энергию света.
Определяя интенсивность процесса фотосинтеза в различных лучах солнечного спектра, К.А. Тимирязев показал, что наиболее интенсивное усвоение углекислого газа наблюдается в красных лучах. Затем по направлению к зеленой части спектра процесс фотосинтеза постепенно ослабевает. В зеленых лучах фотосинтез минимальный. Это и понятно, так как именно зеленые лучи хлорофиллом почти не поглощаются. В сине-фиолетовой части спектра наблюдается второй подъем интенсивности фотосинтеза. Таким образом, если представить себе интенсивность фотосинтеза в виде кривой, то она будет иметь два максимума соответственно двум максимумам поглощения хлорофилла. Ряд пиков интенсивности фотосинтеза соответственно отдельным линиям поглощения хлорофилла не наблюдается, так как хлорофилл в хлоропластах находится в такой концентрации, при которой линии поглощения частично сливаются и образуются два основных максимума. Интенсивность процесса фотосинтеза в различных участках спектра получила название спектра действия. Можно сделать вывод, что спектр поглощения хлорофилла и спектр его действия в процессе фотосинтеза совпадают. Дальнейшие исследования внесли определенные уточнения как в отношении лучей, поглощаемых разными формами хлорофилла, так и в отношении их влияния на процесс фотосинтеза. Однако общие закономерности, установленные К.А. Тимирязевым, остались в силе.
Важное значение имеют исследования К.А. Тимирязева по эффективности использования энергии в красном и сине-фиолетовом участках спектра. Тимирязев провел сравнение интенсивности и эффективности поглощения энергии к разных лучах солнечного спектра. Оказалось, что поглощенная энергия в красном участке спектра используется более полно. Из этого наблюдения К.А. Тимирязев сделал вывод, что поглощенная энергия лучей разного качества, разной длины волны используется в фотохимических реакциях с разной эффективностью. В процессе естественного отбора растения приспособились к поглощению именно тех лучей, энергия которых используется в процессе фотосинтеза наиболее эффективно.
Мысли К.А. Тимирязева получили блестящее подтверждение после того, как А. Эйнштейном была сформулирована теория фотоэффекта. Из теории фотоэффекта следует, что интенсивность любой фотохимической реакции определяется не количеством поглощенной энергии, а числом поглощенных квантов. Между тем, как уже упоминалось, величина квантов в разных лучах солнечного спектра различна. В красных лучах кванты мельче и характеризуются меньшей энергией. По мере того как уменьшается длина волны, растет энергия квантов. В связи с этим на одно и тоже количество поглощенной энергии в красных лучах по сравнению с сине-фиолетовыми приходится большее число квантов и соответственно большее количество прореагировавших молекул в фотохимических реакциях, в том числе и при фотосинтезе. Правда, могут быть кванты, несущие так мало энергии, что ее не хватает на то, чтобы вызвать химический эффект. Иначе говоря, для фотохимических реакций существует нижний предел энергии, т. е. верхний предел длины волны, после которого они неосуществимы. Так, (ротохимические реакции возможны в пределах величины квантов от 147 до 587 кДж/моль. Таким образом, в квантах красного света (176 кДж/моль h\) заключено достаточное количество энергии для осуществления фотохимической реакции. Вместе с тем при поглощении квантов синего света (261 кДж/моль кч) реагирующие молекулы будут получать избыток энергии, который выделяется и виде тепла или света.
Таблица 2 Использование энергии света лучей различной длины при фотосинтезе клетками водоросли хлореллы
| Длина волны, нм | Использование лучистой энергии, в относительных величинах (экспериментальные данные) | Использование лучистой энергии, в относительных величинах, исходя из величины квантов (теоретические данные) |
| 100,0 | 100,0 | |
| 80,0 | 87,5 | |
| 76,0 | 82,5 | |
| 61,5 | 63,0 |
Следовательно, использование энергии зависит от качества света. Это было подтверждено исследованиями О. Варбурга. В этих исследованиях впервые была установлена величина фотосинтетической работы, производимой за счет 1 Дж поглощенной лучистой энергии. Эта величина возрастает по мере увеличения длины волны. Данные таблицы 6 показывают совпадение теоретических расчетов и экспериментально полученных величин.
Таким образом, количество молекул СО2 и Н2О, использованных в процессе фотосинтеза, пропорционально числу поглощенных квантов. Однако число квантов, необходимое для протекания различных фотохимических реакций, неодинаково. Редкая фотохимическая реакция имеет квантовый расход, равный единице. Он может быть значительно больше единицы, так как не все возбужденные молекулы вступают в реакцию. Может быть и меньше единицы, если благодаря цепным взаимодействиям в реакцию вступают не только возбужденные молекулы.
Квантовый расход процесса фотосинтеза, т. е. количество квантов, необходимое для того, чтобы одна молекула СО2 восстановилась до углеводов, окончательно не установлен. Все же большинство исследований показывает, что для восстановления одной молекулы СО2 до углеводов нужно 8—9 квантов света. Противоположной величиной квантовому расходу является квантовый выход — это количество ассимилированного СО2 при поглощении одного кванта. Квантовый выход составляет ‘/g или ‘/9 М. Анализ квантового расхода, наблюдаемого в различных участках солнечного спектра, позволил также доказать роль каротиноидов в процессе фотосинтеза. Исследования А.А. Рихтера, а затем Р. Эмерсона показали, что в той части спектра, где лежит максимум поглощения каротиноидов, т. е. между синими и зелеными лучами, на их долю приходится 70% от всего поглощения и лишь 30% энергии поглощается хлорофиллом. В этой части спектра расход квантов приближается к теоретически возможному, только если принять, что кванты света, поглощенные каротиноидами, передаются хлорофиллу и, таким образом, используются в процессе фотосинтеза. Правда, кванты света, поглощенные каротиноидами, используются менее эффективно по сравнению с квантами, поглощенными непосредственно хлорофиллом.
Фотосинтез — это сложный многоступенчатый окислительно-восстановительный процесс, в котором происходит восстановление углекислого газа до уровня углеводов и окисление воды до кислорода. Фотосинтез включает как световые, так и темновые реакции. Был проведен ряд экспериментов, доказывающих, что и процессе фотосинтеза происходят не только реакции, идущие с использованием энергии света, но и темновые, не требующие непосредственного участия энергии света. Можно привести следующие доказательства существования темновых реакций в процессе фотосинтеза: 1) фотосинтез ускоряется с повышением температуры. Отсюда прямо следует, что какие-то этапы этого процесса непосредственно не связаны с использованием энергии света. Особенно резко зависимость фотосинтеза от температуры проявляется при высоких интенсивностях света. По-видимому, в этом случае скорость фотосинтеза лимитируется именно темновыми реакциями; 2) эффективность использования энергии света в процессе фотосинтеза оказалась выше при прерывистом освещении. При этом для более эффективного использования энергии света длительность темновых промежутков должна значительно превышать длительность световых.
Процесс фотосинтеза включает следующие этапы:
1) фотофизический; 2) фотохимический (световой); 3) ферментативный (темновой).
2.1 Фотофизический этап фотосинтеза
Согласно законам фотохимии, при поглощении кванта света атомом или молекулой какого-либо вещества электрон переходит на другую, более удаленную орбиталь, т. е. на более высокий энергетический уровень (рисунок 1). Наибольшей энергией обладает электрон, отдаленный от ядра атома и находящийся на достаточно большом расстоянии от него. Вместе с тем, чем ближе к ядру, тем меньше энергия электрона. Каждый электрон переходит на более высокий энергетический уровень под влиянием одного кванта света, если энергия этого кванта ровна разнице между этими энергетическими уровнями.
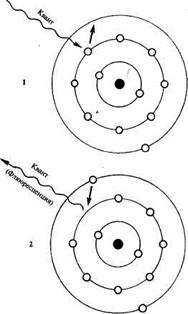
— поглощение фотона, возбуждение атома при переходе электрона;
— возврат электрона выделение энергии в виде излучения.
Все фотосинтезирующие организмы содержат какой-либо тип хлорофилла. Исключением являются бактерии (Halobacterium halobium), содержащие бактериородопсин, с помощью которого они поглощают кванты света.
В молекуле хлорофилла два уровня возбуждения. Именно с этим связано и то, что он имеет две основные линии поглощения. Первый уровень возбуждения обусловлен переходом на более высокий энергетический уровень электрона в системе сопряженных двойных связей, а второй — с возбуждением неспаренных электронов атомов азота и кислорода в порфириновом ядре. При поглощении света электроны переходят в колебательное движение. Наиболее подвижными в молекуле являются делокализованные электроны, орбитали которых как бы размазаны, обобщены между двумя ядрами. Особенно легко возбуждаются электроны сопряженных двойных связей.
Электрон, кроме того, что он находится на определенной орбитали и вращается вокруг ядра, обладает еще спином (вектором магнитного момента) — характеристикой, которую можно трактовать как направление вращения электрона вокруг своей оси. Спин электрона может принимать два значения. Спины двух электронов, находящихся на одной орбитали, противоположны. Когда в молекуле все электроны расположены попарно, их суммарный спин равен нулю. Это основное синглетное состояние (S0). В основном энергетическом состоянии S0 молекула находится
в тепловом равновесии со средой, все электроны попарно занимают орбитали с наименьшей энергией. При поглощении света электроны переходят на следующие орбитали с более высоким энергетическим уровнем. При этом имеются две возможности: если электрон не меняет спина, то это приводит к возникновению первого и второго синглетного состояния (S *1, S*2). Если же один из электронов меняет спин, то такое состояние называют триплетным (Т*1).
Наиболее высокий энергетический уровень — это второй синглетный уровень S*2. Электрон переходит на него под влиянием сине-фиолетовых лучей, кванты которых содержат больше энергии. В первое возбужденное S*l состояние электроны могут переходить, поглощая более мелкие кванты красного света..
Из возбужденного, первого синглетного и триплетного состояния молекула хлорофилла также может переходить в основное. При этом ее дезактивация (потеря энергии) может проходить: 1) путем выделения энергии в виде света (флуоресценция и фосфоресценция) или в виде тепла; 2) путем переноса энергии на другую молекулу пигмента; 3) путем затрачивания энергии на фотохимические процессы (потеря электрона и присоединение его к акцептору (рисунок 2). В любом из указанных случаев молекула пигмента дезактивируется и переходит на основной энергетический уровень. Энергия, испускаемая в виде флуоресценции или в виде теплоты, не может быть использована.
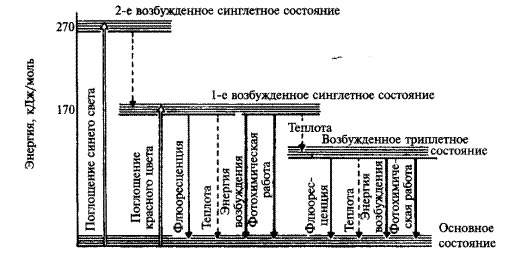
Рисунок 2 – Переходы между возбужденными состояниями хлорофилла после поглощения квантов синего и красного света
В настоящее время показано, что хлорофилл имеет две функции — поглощение и передачу энергии. При этом основная часть молекул хлорофилла — более 90% всего хлорофилла хлоропластов входит в состав светособирающего комплекса (ССК). Светособирающий комплекс выполняет роль антенны, которая эффективно поглощает свет и переносит энергию возбуждения к реакционному центру. Кроме большого числа (до нескольких сотен) молекул хлорофилла ССК содержит каротиноиды, а у некоторых водорослей и цианобактерий фикобилины. Каротиноиды и фикобилины увеличивают эффективность усвоения света за счет того, что они поглощают свет в тех областях спектра, в которых молекулы хлорофилла поглощают свет сравнительно слабо.
Такое устройство позволяет значительно полнее использовать энергию света. Подсчитано, что каждая молекула хлорофилла на прямом солнечном свету поглощает квант света не чаще чем 10 раз в секунду. Между тем скорость последующих реакций фотосинтеза значительно больше. Из сказанного следует, что при непосредственной связи каждой молекулы хлорофилла с последующей реакцией процесс фотосинтеза шел бы прерывисто. Необходимо учитывать также, что использование энергии света в химических реакциях требует большого количества ферментов. Если бы каждая молекула хлорофилла отдавала энергию света непосредственно на фотохимические процессы, то в листе не хватило бы места для размещения всех необходимых для этого ферментных систем. В процессе эволюции в растениях выработался механизм, позволяющий наиболее полно использовать кванты света, падающие на лист подобно каплям дождя. Механизм этот заключается в том, что энергия квантов света улавливается 200—400 молекулами хлорофилла и каротиноидами ССК и как бы стекается к реакционному центру.
В том случае, если молекулы пигментов имеют одинаковый максимум поглощения возможен другой путь передачи энергии, получивший название экситонного. При этом пути электронное возбуждение распространяется в результате межмолекулярной вибрации на комплекс пигментов.
Строение ССК зависит не только от систематического положения фотосинтезирующего организма, но и от условий произрастания. Теневыносливые растения имеют, как правило, больший размер ССК по сравнению с растениями, растущими в условиях высокой освещенности.
Таким образом, в первичных процессах фотосинтеза, связанных с поглощением молекулой хлорофилла кванта света, важную роль играют процессы передачи энергии. Фотофизический этап фотосинтеза и заключается в том, что кванты света поглощаются и переводят молекулы пигментов в возбужденное состояние. Затем эта энергия переносится на реакционный центр, осуществляющий первичные фотохимические реакции: разделение зарядов.
Дальнейшее превращение энергии света в химическую энергию проходит ряд этапов, начиная с окислительно-восстановительных превращений хлорофилла и включая как фотохимические (световые), так и энзиматические (темновые) реакции.
2.2 Фотохимический этап
Происхождение кислорода при фотосинтезе
Большое значение для раскрытия вопроса о сущности фотохимических реакций имело изучение особенностей бактериального фотосинтеза. Впервые на способность бактерий, содержащих пигменты, использовать энергию света для фотосинтеза указал Т. Энгельман (1883). Дальнейшие исследования показали, что окрашенные бактерии содержат пигменты, относящиеся к группе хлорофиллов, а именно бактериохлорофиллы, и синтезируют органическое вещество из неорганических соединений при участии энергии света. Однако этот процесс не сопровождается выделением кислорода. Это связано с тем, что в качестве источника протонов и электронов бактерии используют не воду, а сероводород или другие соединения. Такой тип ассимиляции СО2 получил название бактериального фотосинтеза.
Наиболее распространены содержащие пигменты серные бактерии. Как установил К.Б. ван Ниль, у этих бактерий ассимиляция СО2 сопровождается разложением сероводорода с выделением серы:
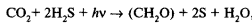 ,
,
где формула (СН2О) означает, что углерод восстановлен до углеводов.
Есть несерные окрашенные бактерии, которые в качестве источника протонов и электронов используют различные органические соединения. Процесс может идти по следующему типу:
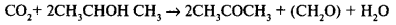
Во всех разобранных случаях происходит восстановление СО2 за счет водорода какого-либо другого соединения. В общей форме это можно представить реакцией:
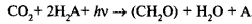 ,
,
где H2A — вещество, которое является источником водорода, а (СН2О) — 1 /6 молекулы сахара. К. Б. ван Ниль высказал предположение, что при фотосинтезе происходит сходный процесс, а именно разложение воды и присоединение водорода к СО2. При этом кислород выделяется. Иначе говоря, процесс фотосинтеза у зеленых растений идет по уравнению:
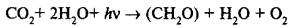
Использование воды в качестве источника водорода дало зеленым растениям в процессе эволюции огромное преимущество в силу повсеместного ее присутствия. Высказанное предположение получило экспериментальное подтверждение в работах академика А.П. Виноградова (1941). Он провел анализ изотопного состава (соотношения 16 О, |7 О, |8 О) кислорода разного происхождения. Оказалось, что кислород, выделенный из воды, воздуха и образующийся прифотосинтезе, имеет одинаковое соотношение изотопов, тогда как кислород СО2 содержит относительно больше тяжелых изотопов. На основании этих исследований было сделано два вывода: 1) в процессе фотосинтеза разлагается вода; 2) в процессе фотосинтеза выделяется кислород, который является основным источником кислорода воздуха.
Поскольку весь кислород фотосинтеза выделяется из воды, общее уравнение фотосинтеза принимает следующий вид:
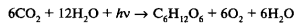
Вода в правой части уравнения не подлежит сокращению, поскольку ее кислород имеет иной изотопный состав (из СО2). Рассмотрение этого уравнения показывает, что фотосинтез — это окислительно-восстановительный процесс, в котором вода окисляется до О2, а углекислый газ восстанавливается до углеводов. Термины «окисление» и «восстановление» являются крайне важными для понимания фотосинтеза. В этой связи необходимо отметить, что окисление — это не только присоединение кислорода, но и отнятие протонов, и потеря электрона, тогда как восстановление — это отнятие кислорода и присоединение протонов или электронов.
В 1937 г. Р. Хилл показал, что изолированные хлоропласты на свету в присутствии какого-либо легко восстанавливающегося вещества (акцептора водорода) окисляют воду. При этом выделяется кислород. В качестве акцептора водорода в опытах Хилла был использован хинон. При этом выделение кислорода хлоропластами на свету протекает в отсутствие углекислого газа (реакция Хилла):
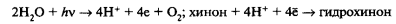
Дальнейшие исследования показали, что те же самые ингибиторы, которые тормозят реакцию Хилла, приостанавливают и выделение кислорода в процессе фотосинтеза. Это дало основание считать, что световая фаза фотосинтеза включает разложение воды. Эти опыты также позволили установить возможность разделения двух процессов: 1) выделение кислорода;
2) восстановление СО2. Таким образом, в процессе фотосинтеза происходит разложение воды, на что затрачивается энергия света.
В 1950 г. было показано, что вместо искусственных акцепторов водорода, примененных Хиллом, можно использовать естественный кофермент никотинами-дадениндинуклеотидфосфат — НАДФ. Изолированные хлоропласты на свету восстанавливают НАДФ, одновременно выделяется кислород. Однако сущность происходящих на свету реакций была выяснена лишь в 1954—1958 гг. благодаря работам Д. Арнона.
2.3 Циклический и нециклический поток электронов. Фотосинтетическое фосфорилирование
Фотохимические реакции фотосинтеза — это реакции, в которых энергия света преобразуется в энергию химических связей, и в первую очередь в энергию фосфорных связей АТФ. Именно АТФ является энергетической валютой клетки, обеспечивающей течение всех процессов. Одновременно под действием света происходит разложение воды, образуется восстановленный НАДФ и выделяется кислород.
Энергия поглощенных квантов света стекается от сотен молекул пигментов ССК к фотохимическому реакционному центру, содержащему особую пару (димер) молекул хлорофилла а, которые характеризуются поглощением в наиболее длинноволновой части солнечного спектра и выполняют роль ловушки энергии возбуждения, блуждающего по пигментам ССК. Наряду с димером хлорофилла в фотосинтетический комплекс входят молекулы первичного и вторичного акцепторов электрона. Молекула хлорофилла, отдавая электрон первичному акцептору, окисляется. Электрон поступает в электронтранспортную цепь.
Совокупность светособирающего комплекса (ССК), фотохимического реакционного центра и связанных с ним молекул — переносчиков электрона составляет фотосистему. По современным представлениям фотосистема является важнейшим структурно-функциональным звеном фотосинтетического аппарата.
В процессе фотосинтеза принимают участие две фотосистемы. Р. Эмерсон показал, что эффективность света с длиной волны 680—700 нм может быть значительно повышена добавлением света с более короткой длиной волны (650—660 нм). Интенсивность фотосинтеза при освещении смешанным светом (с двумя длинами волн) оказалась выше суммы интенсивностей фотосинтеза, наблюдаемой при освещении светом каждой длины волны в отдельности. Это явление называют эффектом усиления или эффектом Эмерсона. Был сделан вывод, что необходимо одновременное световое возбуждение пигментов, различающихся по спектру поглощения. Именно это и привело к понятию о существовании двух фотосистем. Интересно, что эффект усиления наблюдается при изучении фотосинтеза у высших растений и цианобактерий. У других бактерий этот эффект отсутствует и, соответственно, в процессе фотосинтеза участвует одна фотосистема.
Фотосистемы различаются по составу белков, пигментов и оптическим свойствам. Установлено, что фотосистема 1 включает светособирающий комплекс и фотохимический реакционный центр I, в состав которого входит димер хлорофилла, поглощающий свет с длиной волны 700 нм (П700). Фотосистема II включает светособирающий комплекс и фотохимический реакционный центр II, в состав которого входит димер хлорофилла, поглощающий свет с длиной волны 680 нм (П680). Свет поглощается двумя фотосистемами раздельно и нормальное осуществление фотосинтеза требует их одновременного участия.
Перенос по цепи переносчиков включает ряд окислительно-восстановительных реакций. Важно заметить, что при окислительно-восстановительных реакциях происходит перенос либо протонов и электронов, либо только электронов. При этом молекула, отдающая протон или электрон, окисляется, а молекула, воспринимающая протон или электрон, восстанавливается. Различают два типа потока электронов — циклический и нециклический. При циклическом потоке электроны, переданные от молекулы хлорофилла первичному акцептору, возвращаются к ней обратно. При нециклическом потоке происходит фотоокисление воды и передача электрона от воды к НАДФ. Выделяемая в ходе окислительно-восстановительных реакций энергия частично используется на синтез АТФ. Процесс преобразования энергии света в энергию АТФ получил название фотосинтетического фосфорилирования (Д. Арнон), Соответственно двум типам потока электронов различают циклическое и нециклическое фотофосфорилирование.
Общая схема нециклического фотофосфорилирования представлена на рисунке 3. При нециклическом потоке электронов принимают участие две фотосистемы. Как уже упоминалось, энергия квантов света стекается к молекуле пигмента П700, который входит в состав реакционного центра ФСI (фотосистема I). Электрон в молекуле П700 переходит на более высокий энергетический уровень (S*1). В основном состоянии окислительно-восстановительный потенциал П700 составляет +0,43 В. Следовательно, тенденция к потере электрона (окислению) выражена очень слабо. Однако при поглощении кванта света молекула П700 переходит в возбужденное состояние и ее окислительно-восстановительный потенциал изменяется до —0,80 В. Поэтому в возбужденном состоянии молекула П700 легко отдает электрон первичному акцептору (фотохимическая реакция). Энергия света изменяет распределение электронов в кольце молекулы хлорофилла П700 и делает один электрон доступным для переноса. При этом энергия квантов сначала переходит в энергию электронного возбуждения, а затем в энергию разделения зарядов. Так, отдавая электрон, молекула П700 окисляется и остается в виде положительно заряженной молекулы
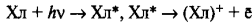
где Хл* — возбужденная форма хлорофилла, (Хл) + — окисленная форма хлорофилла. Электрон с первичного акцептора, которым является мономерная форма хлорофилла а, передвигается через ряд переносчиков, расположенных в порядке возрастания О/В потенциала. Конечным акцептором электрона является НАДФ. Электроны спонтанно текут в сторону менее отрицательного окислительно-восстановительного потенциала. Первичный акцептор, воспринимающий электрон от П700, передает электрон на железосерные белки.
Следующим переносчиком является железосодержащий белок ферредоксин (Е0—0,43). Ферредоксин содержит два атома железа в негеминовой форме. От ферредоксина электрон переносится на НАДФ (Е0-—0,32). Этот перенос осуществляется с помощью специфического белка-фермента (ферредоксин-НАДФ-редуктазы), коферментом которого является флавинадениндинуклеотид (ФАД).
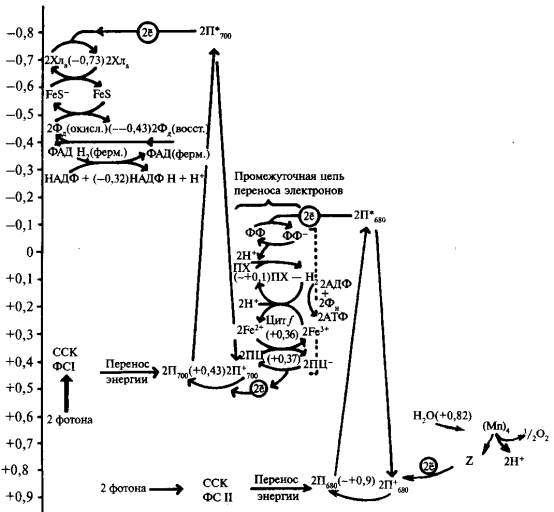
Она ответственна за реакции, связанные с разложением воды и выделением кислорода. Цепь транспорта электронов (ЭТЦ), соединяющая две фотосистемы, в качестве переносчиков электрона включает в себя молекулы пластохинона (ПХ), отдельный электронтранспортный белковый комплекс (так называемый b/f- комплекс) и водорастворимый белок пластоцианин (ПЦ).
В состав реакционного центра фотосистемы II входит хлорофилл а, поглощающий свет с длиной волны — λ680 (П68О). Под влиянием поглощенного кванта света возбужденный электрон от П680 воспринимается первичным акцептором и передается дальше по цепи переносчиков. Образовавшаяся электронная «дырка» в молекуле П680действует как сильный окислитель и через ряд переносчиков при участии ионов марганца и хлора отнимает электрон от воды. Этот электрон заполняет электронную «дырку» в молекуле П680. Происходит фотоокисление воды, и выделяется молекула кислорода:
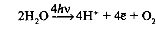
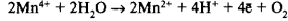
После последовательной передачи четырех электронов от ВРК к П680 + происходит разложение сразу двух молекул воды, сопровождающееся выделением одной молекулы кислорода и четырех ионов водорода, которые попадают во внутритилакоидное пространство хлоропласта.
Перенос электрона по цепи переносчиков от фотосистемы II к фотосистеме I сопровождается образованием АТФ из АДФ и неорганического Фн (АДФ + Фн → АТФ). Суммарное уравнение процесса нециклического фотофосфорилирования может быть выражено следующим образом:
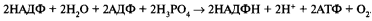
Таким образом, отличительными особенностями нециклического фотосинтетического фосфорилирования являются: 1) окисление двух молекул воды; 2) участие двух фотосистем; 3) передача электронов от молекул воды (первичный донор) через электронтранспортную цепь на НАДФ <конечный акцептор). Продуктами процесса нециклического фотофосфорилирования являются восстановленный никотинамидадениндинуклеотидфосфат (НАДФН + Н + ) и АТФ. Эти соединения в дальнейшем используются в темновой фазе фотосинтеза.
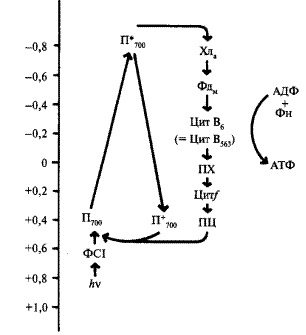
Рисунок 4 – Схема циклического транспорта электронов при фотосинтезе
Общая схема циклического фотофосфорилирования дана на рисунке 4. В этом процессе принимает участие лишь фотосистема I. На схеме показано, что в результате поглощения кванта света молекула П700 отдает возбужденный электрон сначала первичному переносчику, а затем от ферредоксина возвращается к П700. На участке электронтранспортной цепи между цитохромом b и цитохромомƒ образуется АТФ. Таким образом, в этом случае П700 является и донором, и акцептором электрона. Суммарное уравнение циклического фотосинтетического фосфорилирования выглядит следующим образом:
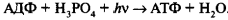
При рассмотрении приведенных схем надо учитывать, что в зависимости от состояния хлоропластов расположение и набор переносчиков в электронтранспортной цепи могут меняться.
Как же осуществляется связь между переносом возбужденного светом электрона по цепи переносчиков и образованием АТФ за счет выделяющейся при этом энергии? Наибольшее признание получила хемиосмотическая теория, разработанная английским биохимиком П. Митчеллом.
Переносчики электронов (электронтранспортная цепь) локализованы в мембранах асимметрично. При этом последовательно чередуются переносчики электронов (цитохромы) с переносчиками электрона и протона (пластохиноны) (рисунок 5).
При поглощении двух квантов света два электрона (поочередно) высвобождаются из П680, пересекают мембрану и замещаются электронами из молекул воды. При этом протоны (из воды) выделяются во внутреннее пространство тилакоида, а атом кислорода диффундирует из хлоропласта. Электроны от П680 передаются на акцептор, расположенный на наружной поверхности мембраны тилакоида. Этим акцептором является пластохинон (ПХ). Пластохинон — производное хинона, в полностью окисленном состоянии содержит два атома кислорода, соединенных с углеродным кольцом двойными связями. В полностью восстановленном состоянии атомы кислорода в бензольном кольце соединяются с протонами:
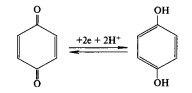
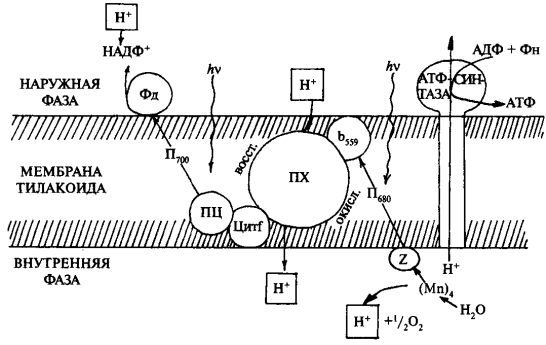
 Рисунок 5 – Схема образования мембранного потенциала и переноса протонов во внутритилакоидное пространство: Фд — ферредоксин; ПЦ — пластоцианин; ПХ — пластохинон; b559, f— цитохромы
Рисунок 5 – Схема образования мембранного потенциала и переноса протонов во внутритилакоидное пространство: Фд — ферредоксин; ПЦ — пластоцианин; ПХ — пластохинон; b559, f— цитохромы
Соответственно и энергия, накапливаемая на мембране в результате концентрирования протонов на одной стороне, имеет две составляющие — химическую и электрическую. Чтобы использовать эту энергию, необходимо обеспечить обратный поток протонов. Энергия, выделяющаяся при этом, и используется для образования АТФ. Накопление протонов внутри тилакоида происходит не только при окислении пластохинона, но и в результате фотоокисления воды.
Сопряжение диффузии протонов через мембрану осуществляется макромолекулярным ферментным комплексом, называемым АТФ-синтазой или сопрягающим фактором (рисунок 6). Этот комплекс по форме напоминает гриб и состоит из двух частей — факторов сопряжения: круглой шляпки F1, выступающей с наружной стороны мембраны (в ней располагается каталитический центр фермента), и ножки F2, погруженной в мембрану. Мембранная часть состоит из полипептидных субъединиц (а, b, с) и формирует в мембране протонный канал, по которому ионы водорода попадают к фактору сопряжения F1. Белок Flпредставляет белковый комплекс, который состоит из девяти субъединиц пяти типов. Белковый комплекс F1 можно сравнительно легко отделить от мембраны, при этом он сохраняет способность катализировать гидролиз АТФ, т. е. является АТФазой.
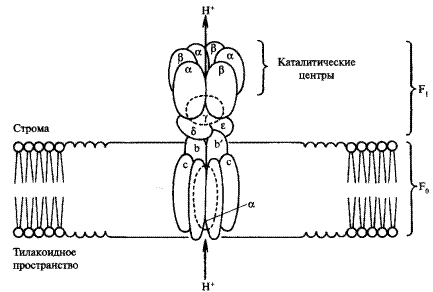
Рисунок 6 – Структура АТФ-синтетазного комплекса
Однако изолированный F1 не способен синтезировать АТФ. Способность синтезировать АТФ — это свойство единого комплекса F0-F1 встроенного в мембрану. Связано это с тем, что работа АТФ-синтазы при синтезе АТФ сопряжена с переносом через нее протонов, путь которых направлен через F0 к F1. Направленный транспорт протонов возможен только в том случае, если АТФ-синтаза встроена в мембрану.
Существуют две гипотезы относительно механизма фосфорилирования (прямой механизм и косвенный) (рисунок 7). Согласно первой гипотезе фосфатная группа и АДФ связываются с ферментом в активном участке комплекса F1. Два протона перемещаются через канал по градиенту концентрации и соединяются с кислородом фосфата, образуя воду. Это делает фосфатную группу весьма реакционноспособной, и она присоединяется к АДФ с образованием АТФ. Согласно второй гипотезе, которая в последние годы получила экспериментальные подтверждения (косвенный механизм), АДФ и Фн соединяются в активном центре фермента (субъединицы α и β фактора F1) спонтанно. Однако образовавшаяся АТФпрочно связана с ферментом, и для ее освобождения требуется энергия. Энергия доставляется протонами, которые, связываясь с ферментом (протонирование), изменяют его конформацию, после чего АТФ высвобождается.
Продукты световой фазы, в которых аккумулирована энергия света, — АТФ и НАДФН Д. Арнон назвал ассимиляционной силой, поскольку они используются в темновую фазу, в ходе которой СО2 восстанавливается до углеводов.